РАЗДЕЛ AMNIOTA. ВЫСШИЕ ПОЗВОНОЧНЫЕ
Класс Reptilia. Пресмыкающиеся.
Пресмыкающиеся, или рептилии, вместе с птицами млекопитающими относятся к высшим позвоночным. От низших позвоночных эти группы отличаются наличием у эмбриона зародышевых оболочек, при помощи которых осуществляется его дыхание и питание.
От земноводных рептилии отличаются следующими основными признаками:
1. Размножением яйцами, имеющими кроме зародышевой оболочки также твердую скорлупу, что в совокупности явилось исключительно важным приспособлением позвоночных к наземной жизни. Из яйца появляется молодое животное, которое отличается от взрослого только размерами.
2. Развитием легочного дыхания, что привело к необходимости защитить голую кожу позвоночных роговым покрытием (роговые чешуи, костные пластинки). Это явилось вторым важным шагом на пути приспособления к сухопутной жизни.
3. Разнообразием условий существования (земля, вода, воздух).
4. Более совершенным кровообращением: в желудочке сердца появилась перегородка (он разделился на правую и левую стороны), а у высших рептилий – архозавров – сердце стало четырехкамерным (у низших рептилий – сердце трехкамерное), и артериальная кровь полностью отделилась от венозной. Развилась вторичная почка - усовершенствовалась выделительная система. В процессе дальнейшего развития центральной нервной системы появилась кора мозга в больших полушариях.
5. Значительными изменениями в строении скелета. В височной области появляются височные впадины, служащие вместилищем челюстной мускулатуры (более сильной, чем у амфибий). Позвоночник приобретает четко выраженную дифференцировку: в частности, обособляется шейный отдел (у земноводных отсутствовал). Это дало животному свободу движений. В теле позвонков постепенно исчезает отверстие для хорды. Парные конечности стали разнообразнее и совершеннее: развились суставные поверхности, конечности удлинились. Это позволило поднять тело высоко, не волоча его по земле, что было крайне необходимым для быстрого передвижения.
Вместе с тем у рептилий сохранились и ряд примитивных признаков: плохо развиты органы чувств (кроме зрения), слуховой аппарат (за небольшим исключением) состоит из одной косточки, т.е. как у амфибий, зубы обычно не дифференцированы и т.д. Как и земноводные, большинство пресмыкающихся являются холоднокровными животными с непостоянной температурой тела. И те, и другие живут обычно в теплых широтах.
Рептилии появляются в середине карбона, в мезозое широко распространены, к концу мезозоя большая часть их вымирает. Современные представители класса – змеи, ящерицы, крокодилы, черепахи, гаттерии и др.
В ископаемом состоянии встречаются как полные скелеты рептилий, так и их части. Довольно часты следы лап рептилий. Их можно найти в местах захоронения скелетов животных, или отдельно. Следы очень разнообразны по облику, размерам и сохранности. Обычны копролиты. Наиболее крупные скопления остатков рептилий наблюдаются в области дельт, т.е. там, где были самые благоприятные условия для захоронения: концентрация трупов течением и их быстрое занесение осадком. В озерах же, наоборот, встречаются обычно отдельные скелеты и их части.
С биологической точки зрения значение пресмыкающихся в истории развития органического мира нашей планеты велико: именно они являются непосредственным промежуточным звеном между амфибиями, птицами и млекопитающими. Родственные связи прослеживаются через котилозавров с амфибиями, через архозавров – с птицами, через зверообразных – с млекопитающими.
Геологическое значение рептилий также велико. Сухопутные рептилии (котилозавры, зверообразные, динозавры) широко используются в стратиграфии верхнепалеозойских и мезозойских, в основном континентальных отложений. Остатки же морских пресмыкающихся (ихтиозавров, плезиозавров, мозазавров) имеют меньшую ценность из-за их редкости, однако при их обнаружении окаменелости применяют в основном для расчленения верхнеюрских и меловых отложений.
Класс Пресмыкающихся объединяет семь подклассов: Cotylosauria – Котилозавры, Chelonia – Черепахи, Ichthyopterygia – Ихтиозавры или Рыбоящеры, Synaptosauria – Синаптозавры, Lepidosauria – Чешуйчатые, Archosauria – Архозавры, Crocodilia – Крокодилы, Synapsida – Зверообразные.
Подкласс Cotylosauria. Котилозавры. C3-T. Это первые настоящие наземные позвоночные, которые дали начало остальным подклассам рептилий (рис.9).
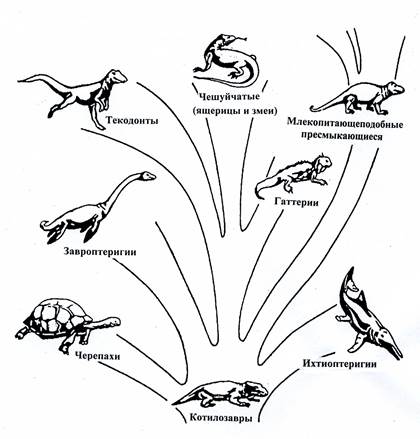
Рис. 9. Схема эволюции пресмыкающихся от котилозавров.
Котилозавры (рис. 10-11) произошли от амфибий – антракозавров, с которыми схожи строением черепа. У них сохранился панцирь на голове. Такой тип черепа из покровных окостенений без височных впадин называется анапсидным. Череп скульптирован. Группа включает как мелких наземных ящерицеобразных животных, так и крупных растительноядных, величиной с бегемота. Морда короткая, реже умеренно удлиненная. Конечности были короткими и массивными. Известный представитель котилозавров - парейазавр (щекастый ящер) (рис.11 б-в, 27) – неуклюжее грузное животное до 3 м длиной. Череп широкий большой скульптированный с массивными выступами («щеками») по бокам, морда округлая, зубы долотовидные. Нередко был спинной панцирь из мозаично расположенных костных пластинок. Растительноядный.
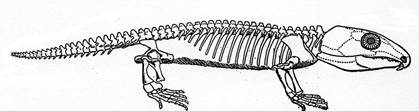
Рис. 10. Класс Рептилии. Котилозавр Labidosaurus (P1).
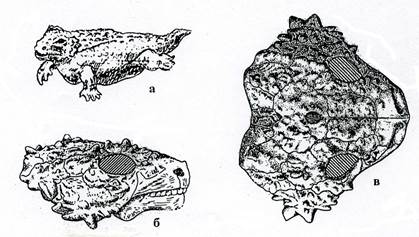
Рис. 11. Класс Рептилии. Котилозавры: а - Scutosaurus (P2), б-в
- парейазавр Nanoparia (P1)(б – череп сбоку, в – череп сверху).
Котилозавры вели различный образ жизни, по способу питания среди них были хищники, растительноядные, моллюскоядные, насекомоядные. Остатки котилозавров многочисленны в пестроцветных фациях дельт и аллювиальных равнин. Животные используются в стратиграфии перми, нижнего и среднего триаса.
Подкласс Chelonia. Черепахи. T3-Q. Черепахи произошли от котилозавров (рис. 11) в перми, но остатки их первых представителей нашли только в верхнем триасе. Эти черепахи уже обладали почти всеми характерными признаками группы.
В подкласс объединяют животных с коротким и широким туловищем, заключенным в костный панцирь. Его и находят в ископаемом состоянии (рис. 12).
Черепахи имели синапсидный тип черепа, т.е. он имел височную впадину, расположенную ниже сочленения чешуйчатой и заднеглазничной костей. Зубы отсутствуют, вместо них имеется роговой клюв. Черепахи, почти не изменившись, дожили до наших дней.
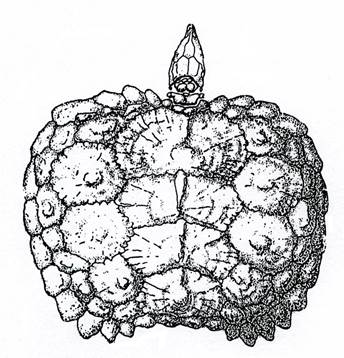
Рис. 12. Класс Рептилии. Черепаха Triassochelys (Т1).
В геологии черепахи используются в основном как показатели климата прошлых эпох. Так, нахождение речных и озерных черепах указывает на влажный и теплый климат. Наличие роющих черепах с уплощенным панцирем свидетельствует об открытых степных или пустынных ландшафтах. Некоторые черепахи имеют значение в стратиграфии палеогена и неогена.
Подкласс Ichthyopterygia. Ихтиозавры, или рыбоящеры. T-K2. Подкласс объединил морских рептилий, утративших связь с сушей. Эти животные имели рыбообразное тело, мягкий спинной плавник и жесткие хвостовые плавники, парные конечности преобразованы в ласты (рис. 13). Длина тела некоторых форм превышала 10 м. Череп с длинной мордой и небольшой височной ямой. Зубы хорошо развиты, они конические, однорядные, лабиринтовые. Небных зубов обычно нет. Посредине хвост в той или иной степени изогнут вниз: у триасовых форм – незначительно, у юрских и меловых - очень резко. В передней конечности 4 или 5 пальцев, в задней – 3 или 4. Чешуйчатый покров не установлен, но в коже была развита система роговых пластин. Ихтиозавры перешли к живорождению, о чем свидетельствуют находки скелетов молодых особей внутри скелетов взрослых.
Ихтиозавры внешне похожи на рыб и дельфинов, к рыбам их первоначально и относили. Многие из ихтиозавров были хищниками и съедали своих же сородичей, часто они питались моллюсками, в том числе белемнитами.

Рис. 13. Класс Рептилии. Ихтиозавр Stenopterygius (J1). Скелет с отпечатком и остатками кожи.
Ихтиозавры известны с верхней части нижнего триаса, их расцвет приходится на раннюю юру, в позднем мелу они вымирают. Ихтиозавры как и другие водные рептилии, имеют меньшую стратиграфическую ценность по сравнению с сухопутными рептилиями из-за их относительной малочисленности, тем не менее они используются для расчленения юрских и меловых отложений.
Ихтиозавры захоронены в сланцах, мергелях, глинах, доломитах совместно с белемнитами, иглокожими, рыбами и др. Многочисленны как находки отдельных костей этих животных, так и целых скелетов, иногда с отпечатками мягких тканей. Скелеты могут быть заключены в известковые конкреции. Основные местонахождения остатков образовались на месте мелководных бухт.
Подкласс Synaptosauria. Синаптозавры. Т-K. Эти пресмыкающиеся вели водный образ жизни. Они имели обычно крупное, широкое (боченковидное) туловище, короткий хвост и часто длинную шею. Длина тела достигала 15 м.
Тип черепа эвриапсидный, характеризующийся одной верхней височной ямой. Конечности – ластовидные.
Широким распространением из синаптозавров пользовались завроптеригии (плезиозавры, плиозавры) и плакодонты.
Плезиозавры (рис. 14) – животные с маленькой головой и длинной, гибкой шеей. Обитали вблизи берега, питались рыбой. Классические местонахождения плезиозавров находятся в Англии.

Рис. 14. Класс Рептилии. Плезиозавр Muraenosaurus (J3).
Плиозавры (рис. 15)– хищники огромных размеров с короткой шеей и крупной головой (череп до 4 м в длину), с сильными коническими зубами. Обитатели открытых морей. Принадлежали к числу опаснейших морских хищников своего времени.
Завроптеригии входят в состав типовых комплексов позвоночных верхнего триаса-нижнего мела и используются для расчленения и определения возраста отложений этого уровня.
Плакодонты (рис. 16) похожи на черепах, питались моллюсками.
Есть мнение, что завроптеригии и плакодонты возникли от котилозавров. Завроптеригии появились в триасе, вымерли в конце мела. Плакодонты жили только в триасе. Согласно другой точке зрения синаптозавры берут свое начало от примитивных диапсид, вероятно, близким эозухиям (см. ниже).
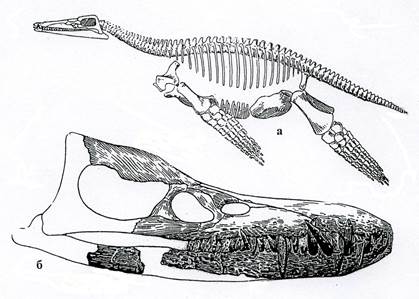
Рис. 15. Класс Рептилии. Плиозавры: а – Peloneustes (J3), б – череп Pliosaurus сбоку (J3).
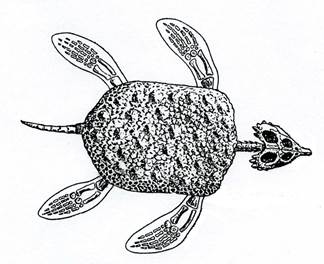
Рис. 16. Класс Рептилии. Плакодонт Placochelys (T3).
Остатки синаптозавров встречаются редко, поэтому стратиграфическое значение их невелико. Но плакодонты, имеющие узкое распространение, используются как руководящие формы для триаса.
Подкласс Lepidosauria. Чешуйчатые. Р2-Q. Это примитивные ящерицеобразные пресмыкающиеся с двумя височными дугами, обладающие пластинчатыми поясами конечностей. Парные конечности в основном умеренной длины, но у змей и некоторых ящериц они исчезают, хотя рудименты поясов конечностей могут сохраняться. У некоторых водных форм конечности становятся ластовидными. У лепидозавров сохраняется обычный для ранних четвероногих примитивный тип передвижения с волнообразным изгибанием туловища.
Вымершие чешуйчатые представлены наземными ящерицеобразными эозухиями и хищными морскими ящерицами – мозозаврами. Они имеют диапсидный тип черепа и являются переходной группой между примитивными рептилиями – котилозаврами и прогрессивными рептилиями – архозаврами.
Эозухии (рис. 17) в большинстве имели некрупные размеры, они появились в перми, в триасе дали начало другим чешуйчатым: клювоголовым и ящерицам.
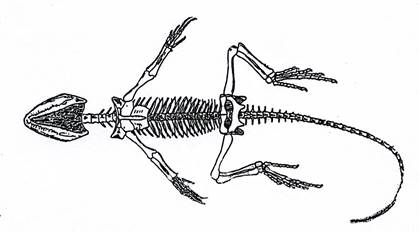
Рис. 17. Класс Рептилии. Эозухия Homoeosaurus (J3).
Мозозавры (рис. 18) – морские ящерицы, жившие в позднем мелу, имели удлиненное тело до 15 м в длину и две пары пятипалых конечностей – ласт, хвост превращен в плавник, который являлся главным органом движения.
| |
| Рис. 18. Класс Рептилии. Мозозавры – морские ящерицы (К2): а – скелет, б-в – реконструкция животных. | 
|
В конце мела от ящериц обособились змеи. Современные представители подкласса – змеи, ящерицы, хамелеоны, гаттерии. Гаттерии – единственный современный вид из клювоголовых ящерицеобразных, живет в Новой Зеландии.
Лепидозавры жили в разных услових (морях, болотах, озерах, на суше и т.д.), поэтому их остатки встречают в разных породах. Стратиграфическое значение лепидозавров невелико. Но те из них, которые имеют узкий диапазон распространения, например, мозозавры, с успехом используются для расчленения мезозойских отложения.
Подкласс Archosauria. Архозавры (древние ящерицы). Р2-Q. Самая многочисленная группа рептилий с диапсидным типом (две височные впадины и часто предглазничное отверстие) черепа. По способу размножения архозавры были яйцекладущими животными, но возможно некоторые формы, живущие в воде, были живородящими. Архозавры произошли от чешуйчатых – эозухий. Первые архозавры, жившие в перми и триасе, объединены в надотряд текодонтов.
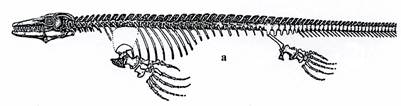
Текодонты имели длинный узкий череп и текодонтные зубы. Это зубы, которые расположены в особых ячейках по краям челюстей. Передние конечности у текодонтов были короче, чем задние. Все они являлись хищниками. Текодонты разделяются на фитозавров (рис. 19) – крокодилообразных текодонтов с удлиненным рылом и кожным панцирем, живших в пресноводных бассейнах триаса, и псевдозухий (рис. 20, внизу) - некрупных подвижных наземных форм с панцирем или без него, распространенных в перми и триасе.
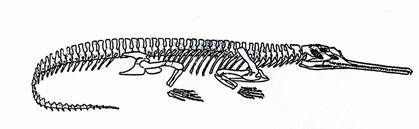
Рис. 19. Класс Рептилии. Скелет фитозавра Belodon (T).
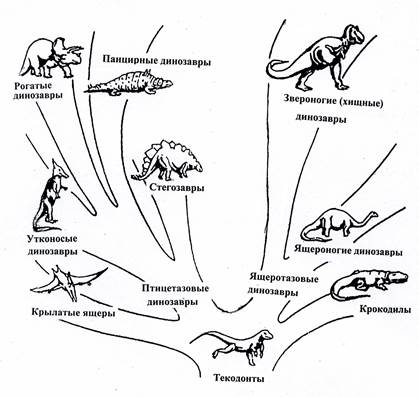
Рис. 20. Схема эволюции пресмыкающихся от текодонтов.
Текодонты дали начало всем остальным группам этого подкласса: разнообразным динозаврам, летающим ящерам и крокодилам.
Представители надотряда используются в стратиграфии в основном триасовых отложений.
Надотряд динозавров. Dinosauria (страшная ящерица). Т2-К2. Это – самая многочисленная, господствующая в мезозое, группа рептилий. Объединяет разнообразных по морфологии, размерам и образу жизни животных. Здесь есть небольшие рептилии величиной с кошку и крупные - до 30 м в длину, двуногие и четвероногие, хищники и питавшиеся растениями, несшие костные панцири и «голые».
Выделяются два отряда динозавров: ящеротазовые и птицетазовые.
Отряд Ящеротазовые динозавры. Делятся на две группы: звероногие (тероподы) и ящероногие (зауроподы). Появились в триасе, расцвет группы приходится на юру и ранний мел, в конце мела вымирают.
| Т е р о п о д ы объединили двуногих животных с укороченны-ми передними конечностями, которые передвигались на задних лапах, опираясь на сильный хвост. Среди них были хищники и растительноядные. Размеры достигали 14 м в длину. Некоторые тероподы имели перьевой покров. Характерный представитель группы – тираннозавр (рис. 21). Рис. 21. Класс Рептилии. Ящеротазовый динозавр из группы звероногих – Tirannosaurus. Реконструкция внешнего вида | 
|
З а у р о п о д ы (рис. 22) относились к растительноядным организмам. Они передвигались на четырех лапах. Размеры их головного мозгабыли очень маленькими. Но это были самые крупные животные, жившие кода-либо на нашей планете. Так, диплодоки и бронтозавры имели длину тела до 30 м и вес до 30 тонн. Считается, что они вели полуназемный образ жизни и часть жизни проводили в воде. По-видимому, они были отличными пловцами, причем главным органом плавания являлся хвост.

Рис. 22. Класс Рептилии. Ящероногий динозавр Diplodocus (J3) - диплодок.
Отряд Птицетазовые динозавры. Все рептилии этого отряда являются растительноядными. Они имели разные размеры и морфологию, многие имели защитные приспособления: панцирь, рога, щиты и др. Появляются в начале мезозоя, а к концу мела вымирают. Выделяются четыре группы этих динозавров: птиценогие динозавры, стегозавры, панцирные и рогатые динозавры.
Птиценогие динозавры ходили на двух задних конечностях. Они жили на суше и в неглубоких водоемах. Один из их представителей, игуанодон (рис. 23), представлял собой крупное животное с длиной тела до 5 м и черепом похожим на лошадиный. Зубы напоминали ряд зубов млекопитающих. Передние конечности игуанодонов были короткими, пятипалыми, задние конечности были трехпалые с копытами.
Стегозавры (рис. 24) первоначально передвигались на задних конечностях, а позже перешли к четвероногому хождению. Они имели большое туловище, маленькую голову и крупный хвост. Длина тела до 6 м. В качестве защитного приспособления на его спине имелись костные пластинки размером до 1 м, а на конце хвоста – острые шипы. Распространение стегозавров – юра-ранний мел.
Панцирные динозавры (анкилозавры) (рис. 25) были крупными животными с широким уплощенным телом (длина до 5 м), покрытым массивным панцирем из костных пластин. Стратиграфическое распространение их достаточно узкое – мел.
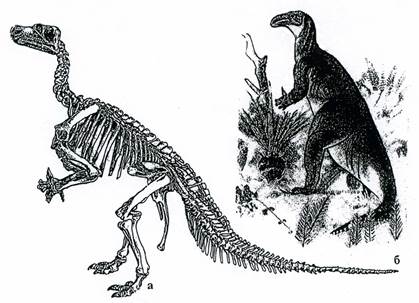
Рис. 23. Класс Рептилии. Птиценогий динозавр Iguanodon (K1) – игуанодон:а – скелет, б – реконструкция животного.
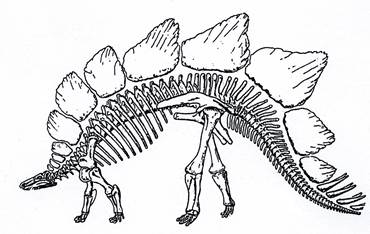
Рис. 24. Класс Рептилии. Птицетазовый динозавр Stegosaurus (J3) – стегозавр.

Рис. 25. Класс Рептилии. Панцирный динозавр – Ankylosaurus (K2) – анкилозавр.
В позднем мелу обитали рогатые динозавры (цератопсы) – крупные (до 7 м в длину) грузные рептилии, ходившие на четырех ногах. Характерная их особенность – наличие на черепе костного «воротника», который защищал область шеи. На конце морды и над глазами располагались рога. Типичный представитель группы – трицератопс (рис. 26).

Рис. 26. Класс Рептилии. Рогатый динозавр Triceratops (K2): а – скелет, б – реконструкция.
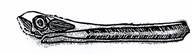
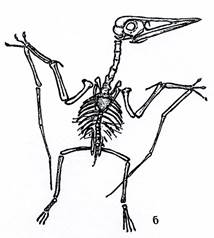
Надотряд Pterosauria. Птерозавры (крылатые ящеры) – летающие животные, внешне напоминающие птиц (рис. 27-28).

Рис. 27. Класс Рептилии. Птерозавры, или крылатые ящеры: а – длиннохвостый птерозавр Rhamphorhinchus (J) – рамфоринхус, б – гигантский короткохвостый птерозавр Pteranodon (K) – птеранодон.
| 
|
| Рис. 28. Класс Рептилии. Крылатый короткохвостый ящер птеродактиль (J): а – череп с многочисленными узкими зубами, б – скелет, длина – 12 см. |
Четырехпалые с сильно удлиненным четвертым пальцем передние конечности птерозавров были преобразованы в крылья. Между конечностями и боками тела была натянута летательная перепонка. Челюсти – вытянутые, снабженные обычно зубами. Хвосты - разной длины.
Птерозавры произошли от текодонтов. Жили с позднего триаса до конца мела.
Характерными представителями надотряда являются рамфоринхусы,птеродактили и птеранодоны.
Первые были размером с небольшую птицу, имели зубы и длинный хвост, заканчивающийся кисточкой.
Размеры птеродактилей - также небольшие, их клюв был снабжен зубами, но хвост был коротким или отсутствовал.
Птеронодоны являлись беззубыми ящерами с коротким хвостом и высоким гребнем в верхней части черепа, имели большие размеры (размах крыльев достигал 8 м).
У некоторых птерозавров обнаружен шерстный покров.
Стратиграфическое значение птерозавров невелико.
Подкласс Crocodilia. Крокодилы.Т2-ныне. (рис. 29). Тело этих рептилий покрыто роговыми щитками, под которыми расположены костные пластинки. Туловище приплюснуто, хвост – длинный, задние конечности длиннее передних.
Появились крокодилы в триасе, являются самыми организованными из современных пресмыкающихся. Сейчас населяют пресные и опресненные водоемы тропиков. Все крокодилы – хищники. Морские крокодилы используются в стратиграфии юры, пресноводные – верхнего мела.

Рис. 29. Класс Рептилии. Морской крокодил (MZ).
Подкласс Synapsida. Зверообразные. С3-К1. Эти рептилии имели синапсидный тип черепа. В ходе эволюции приобрели черты млекопитающих: увеличилась зубная кость нижней челюсти, появилась дифференциация зубов и т.д. Большинство синапсид - хищники.
В конце карбона они возникли от котилозавров, в свою очередь, их представители в конце триаса дали начало млекопитающим. Есть мнение, что зверообразные не относятся к настоящим рептилиям, а образуют самостоятельный класс – промежуточный между рептилиями и млекопитающими. Примером зверообразных является Inostrancevia (рис. 30, 31). Рептилия имела дифференцированные зубы: резцы, заклыковые зубы, мощные клыки, а на пальцах – большие когти. Животное достигало 3 м в длину при длине черепа 50 см.

Рис. 30. Класс Рептилии. Зверообразный хищник Inostrancevia (P2) (сверху), котилозавр Pareiasaurus (P2) (внизу).
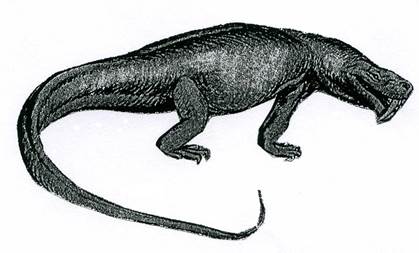
Рис. 31. Класс Рептилии. Реконструкция представителя подкласса
зверообразных – Inostrancevia (P2).
Класс Aves. Птицы.
Птицы в современной фауне представляют наиболее (после рыб) многочисленный класс позвоночных. Они хорошо приспособились к жизни в воздушной среде. Произошли от динозавров.
Основные признаки птиц и их отличия от пресмыкающихся:
- передние конечности птиц превращены в крылья, которые служат для летания и не приспособлены к другим функциям, которые обычны для передних конечностей других животных,
- теплокровны, поддерживают постоянную температуру тела,
- тело покрыто перьями, которые защищают от холода и помогают при полете,
- кровеносная, нервная системы и органы чувств более совершенны; сердце четырехкамерное,
- головной мозг больших размеров,
- скелет прочный и легкий, многие элементы скелета полые, содержащие в своих полостях воздух.
Часть из перечисленных признаков встречаются и у рептилий, но в целом для них не характерны. Например, перьевой покров был у некоторых динозавров.
В классе птиц выделено три подкласса: Ящерохвостые птицы, Зубастые птицы и Веерохвостые (или Новые) птицы.
Ящерохвостые птицы (J3). Самые древние птицы, сочетающие в своем строении признаки пресмыкающихся и более поздних птиц. Некоторые из них относятся к роду Archaeopteryx (рис. 32).
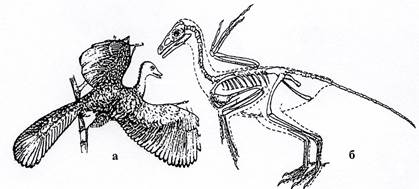
Рис. 32. Класс Птицы. Ящерохвостая птица Archaeopteryx (J3):
а – общий вид, б – скелет.
Эта птица была величиной с ворону. Тело покрыто перьями. Череп был маленьким и плоским, что обычно для пресмыкающихся. Кости черепа сращены, большие полушария головного мозга были совершенно гладкие (установлено по слепкам мозговой коробки). Клюв отсутствовал. Челюсти были усажены маленькими острыми зубами, сидящими в ячейках. Позвоночник и многие кости скелета не сращены и не полые. Передние конечности представляли собой крылья с тремя свободными пальцами с когтями. Хвост состоял из свободных позвонков. Эти птицы плохо летали. Жили в лесах. Предками настоящих птиц не являются (тупиковая ветвь).
Зубастые птицы (К). По строению близки современным птицам, но имели зубы. В эту группу входят гесперорнисы и ихтиорнисы.
Гесперорнисы (рис. 33 а)– крупные до 180 см в длину, водоплавающие птицы, рыбоядные, похожи на гагар. У них был редуцирован плечевой пояс, исчезли кости крыла. Ноги расходились в стороны под углом и работали как весла. Эта птица была приспособлена только к плаванию, способность к полету полностью утратила.
Ихтиорнисы (рис.33 б-в)– до 25 см в длину, летающие, похожи на чаек.
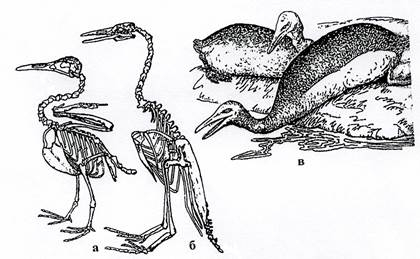
Рис. 33. Класс Птицы. Зубастые птицы: а – Hesperornis (K2), б-в – Ichthyornis (K2)(б – скелет, в – реконструкция).
Веерохвостые (или новые) птицы. Это большая групп ископаемых и современных птиц. Среди них есть бегающие (страусы), плавающие (пингвины) и летающие (большинство) семейства птиц. Летающие веерохвостые птицы известны с позднего мела.
Остатки птиц находят редко, так как их кости быстро разрушаются. Иногда в ископаемом состоянии сохраняются перья или скорлупа.
Стратиграфическое значение ископаемых птиц невелико.
Класс Mammalia. Млекопитающие.
Млекопитающие представляют собой класс животных, наиболее характерный для кайнозоя. Быстрое появление и развитие отдельных семейств класса типичны для последних 70 млн. лет жизни на Земле.
Первые находки млекопитающих известны из отложений верхнего триаса (рис. 34), в юре и мелу они еще немногочисленны. Произошли от звероподобных рептилий – териодонтов.
Млекопитающие приспособились к различным внешним условиям. Облик их разнообразен. Различны и способы питания, среди них есть растительноядные, плотоядные и всеядные. Прекрасно развиты дыхательная, пищеварительная, кровеносная, выделительная и нервная системы. Конечности пятипалые, но у некоторых животных в связи с образом жизни они видоизменяются. Зубы дифференцированы на резцы, клыки и щечные. Внешние покровы разнообразные: ногти, когти, копыта, роговые чешуи, рога. Тело голое или покрыто волосами. Млекопитающие (кроме первозверей) живородящие, теплокровные. У них развита забота о потомстве, детеныши вскармливаются молоком матери.
В составе класса выделяют шесть подклассов, три из которых, появившиеся с триаса, примитивные по строению, объединены в группу архаичных млекопитающих. Они имели небольшие размеры (как мыши), в конце мезозоя вымерли. Остальные три подкласса появились в мелу и дожили до наших дней.
Первозвери. Это примитивные млекопитающие, имеющие некоторые признаки рептилий. Размножаются яйцами. У некоторых есть зубы (молодые утконосы). Современные представители: утконос, ехидна и проехидна. Произошли от архаичных млекопитающих.
Низшие звери (Сумчатые). У них отсутствует плацента, детеныши рождаются недоразвитыми и очень маленьких размеров. Произошли от архаичных млекопитающих. Современные представители: кенгуру, сумчатые волки и др. Ископаемый сумчатый волк изображен на рис. 35.

| 
|
| Рис. 34. Класс Млекопитающие Древнейшее из известных млекопитающих из верхнего триаса Megazostrodon | Рис. 35. Класс Млекопитающие. Низшие звери. Сумчатый волк Prothylacynus (KZ) |
Высшие звери (Плацентарные). Для них характерно наличие плаценты, благодаря такому приспособлению детеныши рождаются достаточно развитыми и способными сосать молоко. Высоко развит мозг. В подклассе выделяется 27 отрядов, из них 10 – вымершие.
Наиболее примитивные плацентарные – отряд насекомоядные, они явились исходной группой, от которой произошли другие отряды плацентарных.
Среди плацентарных, в частности, имеются следующие отряды: Грызуны (рис. 36 а) (зайцы), Хищные (рис. 36 б, 37 а) (гиены, кошки), Китообразные (дельфины), Древние копытные (рис. 37 б) (фенакодус), Непарнокопытные (рис. 38) (носороги, лошади), Парнокопытные (рис. 39) (олени, верблюды), Хоботные (рис. 40) (мамонт), Приматы (обезьяны, люди).

Рис. 36. Класс Млекопитающие. Высшие звери: а – зайцеобразный грызун Eurymylus (Р),б – гиеновидный хищник Нуaenictitherium (N).
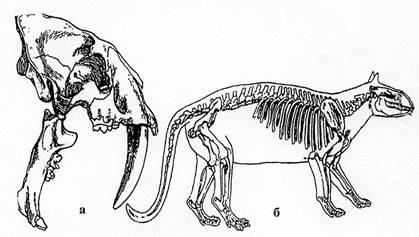
Рис. 37. Класс Млекопитающие. Высшие звери: а – отряд Хищные, череп Machairodus (N2),б – отряд Древние копытные, Phenacodus (Р2).
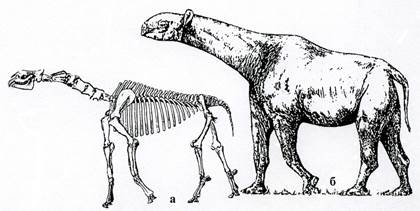
Рис. 38. Класс Млекопитающие. Высшие звери. Отряд непарнокопытные: гигантский носорог Indricotherium (Р3): а – скелет, б – реконструкция.

Рис. 39. Класс Млекопитающие. Рис. 40. Класс Млекопита-
Отряд Непарнокопытные. Больше- ющие. Отряд Хоботные: а - ма-
рогий олень Megaloceras (Q1). монт Mammutus (Q2), б – масто-
дон Mastodon (N-Q2).
Отряд Приматы. Для приматов характерны пятипалые конечности, выполняющие хватательные движения, развитые большие полушарии мозга.
Первые приматы известны с позднего мела. Они близки к насекомоядным.
Человек принадлежит к семейству людей, в составе которого имеется и Homo sapiens L. – человек разумный. Представителей людей делят на древнейших (питекантропов), древних (палеоантропов или неандертальцов) и современных (неоантропов). Существуют разные классификации гоминид. Люди предположительно произошли от человекообразных обезьян. Известны с верхней части неогена (плиоцен).
Полные скелеты млекопитающих находят редко. Преимущественно встречаются такие разрозненные элементы скелета, как челюсти, зубы, рога, ключицы, конечности, части черепа, позвонки, ребра. Нередко на месте необходимо применение специальных методов препарирования, чтобы предупредить разрушение скелетных остатков при высыхании.
Остатки млекопитающих находят в различных породах. Так, довольно часто они встречаются в ледниковых отложениях (например, во флювиогляциальных отложениях террас), в торфах, в отложениях пещер, в вечной мерзлоте и т.д.
Млекопитающие широко используются для расчленения кайнозоя, особенно, четвертичного периода. Это объясняется тем, что остатки их в кайнозое многочисленны, а скорость эволюции очень высокая. Например, в позднем кайнозое (с позднего олигоцена по голоцен) Европы выделено 24 зоны млекопитающих, каждая из которых имеет свой характерный набор животных.
ЗАКЛЮЧЕНИЕ
В органическом мире позвоночные занимают особое место. Морфологические особенности их уникальны и по сравнению с беспозвоночными они являются более совершенными представителями животных.
Количество позвоночных в целом невелико (всего 45 тыс. современных видов), но они весьма разнообразны по приспособительным и жизненным формам. Это связано как с общим высоким уровнем их развития и сложностью строения, так и со способностью приспосабливаться к различным условиям обитания. Позвоночные играют важную роль в биосферных процессах, т.к. обычно завершают трофические связи в биоценозах. Они имеют меньшее стратиграфическое значение по сравнению с беспозвоночными, тем не менее, используются для расчленения, корреляции и определения возраста отложений палеозоя, мезозоя и кайнозоя.
ЛИТЕРАТУРА
1. Еськов К.Ю. История Земли и жизни на ней: учебное пособие. – М.: МИРОС, 2000. – 349 с.
2. Основы палеонтологии. Бесчелюстные. Рыбы / под ред. Д.В.Обручева. – М.: Наука, 1964. – 522 с.
3. Основы палеонтологии. Земноводные, пресмыкающиеся и птицы / под ред. Ю.А.Орлова. – М.: Наука, 1964. – 722 с.
4. Основы палеонтологии. Млекопитающие / под ред. В.И.Громова. – М.: Наука, 1962. – 421 с.
5. Парфенова М.Д. Историческая геология с основами палеонтологии: учебное пособие. – Томск: Изд-во ТПУ, 1998. – 550 с.
6. Шпанский А.В. Основы палеонтологии тетрапод: учебное пособие. – Томск: Изд-во Томск. гос. ун-та систем упр. и радиоэлектроники, 2005. – 216 с.
Дата добавления: 2016-08-07; просмотров: 1558;