Ионный канал как динамическая структура
Если один из участков связывания в канале заполнен, то поступление в свободный участок второго иона будет затруднено в связи с необходимостью дополнительных затрат на преодоление энергии кулоновского взаимодействия с уже находящимся в канале ионом. Попадание иона в левую потенциальную яму приведет к повышению энергетического барьера, соответствующего переходу иона из раствора в правую потенциальную яму (Рис ниже)
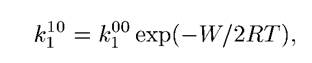
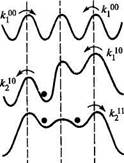
при учете кулоновских взаимодействий вид энергетического профиля зависит от положения и числа ионов в канале. В связи с изменением высоты барьеров изменяются и константы скорости перехода через отдельные барьеры.
константа скорости, соответствующая поступлению второго иона в канал, будет равна (ФОР 6)
 где W — энергия кулоновского взаимодействия: расстояние между потенциальными ямами
где W — энергия кулоновского взаимодействия: расстояние между потенциальными ямами
 *1оа=*1 (РИС 2)
*1оа=*1 (РИС 2)
Зависимость энергетического профиля от заполнения канала при учете кулоновских взаимодействий
fel — константы поступления иона в раствор, &2 — константы выхода иона из раствора. Верхние индексы соответствуют разным состояниям канала с разным заполнением участков связывания
При изменении числа частиц в канале энергетические профили изменяются таким образом, что вход второго иона в канал по сравнению с входом первого затрудняется, а выход второго иона из канала, напротив, идет очень легко. Подробный анализ модели показывает, что кулоновское ионное взаимодействие значительно ускоряет общий ионный транспорт через канал.
Высота барьеров может претерпевать динамические изменения не только за счет кулоновских взаимодействий ионов в канале, но и в результате конформационных перестроек белка, формирующего ионный канал, которые могут инициироваться перескоками иона между отдельными участками связывания.Сильное электрическое поле иона, попадающего в канал, вызывает поляризацию и переориентацию близлежащих полярных групп.В результате появления такого конформационно неравновесного состояния смещается уровень потенциальной энергии в
участке связывания и изменяется высота ближайших барьеров Особенно важное значение могут иметь циклические изменения конформации мембранных белков, обусловленные биоэнергетическими процессами. Такие периодические конформационные перестройки протекают, по-видимому, в АТФазе энер-госопрягающих мембран в процессе ее функционирования, в цитохромах при изменении их окислительно-восстановительного состояния, в бактериородопсине гало-фильных бактерий при возбуждении хромофорной группы. Это обстоятельство послужило основой для гипотезы о том, что системы активного транспорта ионов, использующие энергию света, АТФ или разности редокс-по-тенциалов, представляют собой трансмембранный ионный канал с регулируемой высотой энергетических барьеров
Флуктуации проводимости мембраны.
Электрические флуктуации в мембранах обусловлены изменением числа от
крытых каналов вследствие случайных процессов их формирования и распада.
Анализ флуктуации проводимости позволяет оценить две основные характеристики
канала: среднее время жизни канала в открытом состоянии и проводимость оди
ночного канала.
Суммарная проводимость мембрана определяется статистическим наложением проводимости отдельных каналов, каждый из которые может находиться с определенной вероятностью в закрытом или открытом состояниях.
Переход канала между открытым О и закрытым R состояниями представляет собой в кинетическом отношении реакцию первого порядка с вероятностями закрытия q и открытия р в единицу времени. Среднее статистическое время пребывания одиночного канала в открытом состоянии то обратно пропорционально вероятности его перехода в закрытое состояние q, T0= 1/q
(ФОР 7)
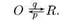
|
 |
Достижение равновесного распределения между открытыми по и закрытыми пR каналами в макроскопической системе с большим числом каналов происходит согласно уравнению кинетики (ФОР 8)
по экспоненциальному закону
nj(y)=n(с чертой)o-n(с чертой)o-n0o – exp[-(p+q)t]
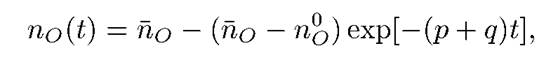 |
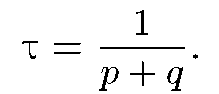
где по — равновесное, по 0(которое с О и 0)— начальное значения величины по] среднее время достижения равновесных значений п с - о и пR в макроскопической системе составляет ти=1/(p+q)
При обычных температурах происходят флуктуации числа открытых и закрытых каналов около их равновесных значений, соответствующие флуктуациям проводимости мембраны.
небольшие спонтанные флуктуации будут затухать с характерным временем
т = 1/(р + q).
|
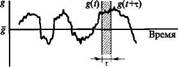
РИС 3
Изменение проводимости мембраны как функция времени
Если значение проводимости мембраны в момент t равно g(t), то через небольшой промежуток времени значение проводимости ^(£ + т) будет иметь близкую величину; иначе
говоря, величина в момент времени (t + т) будет «коррелировать» с величиной в момент времени t. Однако через достаточно большой промежуток времени уже невозможно предсказать величину g(t + x), основываясь на ее первоначальной величине g(t), т.е. g(t + т) и g(t) становятся полностью независимыми
Причина флуктуации состоит в том, что макроскопический ток через мембрану есть результат наложения микроскопических токов, идущих через одиночные каналы, которые ведут себя стохастически и открываются и закрываются с определенной вероятностью.
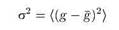 |
Если каналы в мембране открываются и закрываются независимо, то изменения проводимости мембраны представляют собой стохастический процесс. Значения проводимости g колеблются около некоторого среднего значения g с дисперсией а2 или среднеквадратичным отклонением :
значение проводимости флуктуирует вокруг нуля (т.е. проведено вычитание среднего значения). Значение автокорреляционной функции при х — 0, по определению, равно дисперсии флуктуирующего сигнала а2
Дата добавления: 2016-04-11; просмотров: 690;