ПРИНЦИПИАЛЬНЫЕ СХЕМЫ ОСНОВНЫХ ЭЛЕМЕНТОВ СХЕМЫ ТЕРМОРЕГУЛИРОВАНИЯ
Перейдем к описанию принципиальных схем основных элементов схемы терморегулирования (рис. 3.35—3.37).
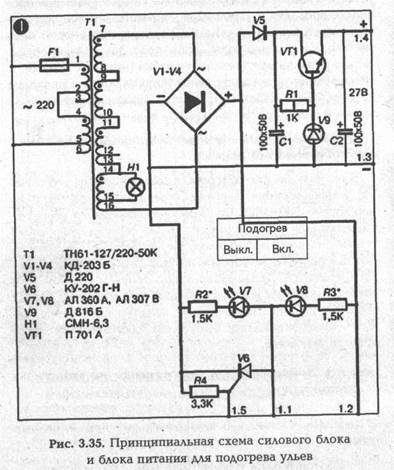
► Силовой блок и блок питания для подогрева ульев
В качестве силового трансформатора Т1 в схеме используется серийный накальный трансформатор типа ТН61-127/220-50К, но может использоваться и любой другой на
|
мощность не менее 200 Вт (для 10-ти ульев). Напряжение вторичной обмотки должно соответствовать выбранному напряжению для подогрева. При этом надо иметь в виду следующее:
1. Реально к подогревателям ЕК будет прикладываться напряжение на 1,5 В ниже расчетного за счет падения этого напряжения на открытом тиристоре V6. В моей схеме к подогревателю реально прикладывается напряжение 22,5 В.
2. За счет естественного повышения величины сопротивления нагревательного элемента при его разогреве ток через подогреватель уменьшается. Реальный ток через один подогреватель ЕК составляет 0,7 А против 0,96 А по расчету.
За счет этих двух причин реальная мощность подогревателя составляет 16 Вт против 23 Вт по расчету.
Эти особенности работы схемы надо обязательно учитывать при расчете реальной мощности подогревателя.
Если мощность изготовленного подогревателя окажется недостаточной, то ее можно увеличить или за счет увеличения питающего'напряжения, снимаемого с трансформатора, или за счет уменьшения сопротивления (длины) нагревательного элемента. В нашем варианте для увеличения реальной мощности нагревателя до 20 Вт надо или увеличить снимаемое со вторичной обмотки питающее напряжение до 30 В (если такая возможность есть), или отрезать около 5 Ом сопротивления, измеренного на «холодной» проволоке.
И еще одно замечание. Подогреватель обеспечивает заданную мощность при номинальном значении питающего напряжения сети в 220 В. Однако, по ряду причин, особенно в сельской местности, напряжение в сети может поддерживаться ниже номинала. В отдельных местностях напряжение сети бывает даже менее 200 В. Поэтому, если окажется, что не хватает мощности подогревателя, поиск причины надо начинать с измерения номинала питающего напряжения сети.
В схеме в качестве выпрямителя используется мостовая схема на диодах V1—V4 типа КД 203Б. Однако могут использоваться и другие силовые диоды на ток 5—10 А. На транзисторе VT1 собрана схема стабилизации для питания микросхемы. Исполнительным элементом схемы терморегулирования является тиристор V6 типа КУ 202 с любым буквенным индексом от Г до Н. Диод V5 выполняет функцию развязки напряжения, прикладываемого к тиристору и подогревателям. Диод V5 может быть любого типа на ток не менее 0,1 А. Лампа Н1 и светодиоды V7, V8 используются для контроля работы схемы терморегулирования.
► Силовой блок для обогрева помещения
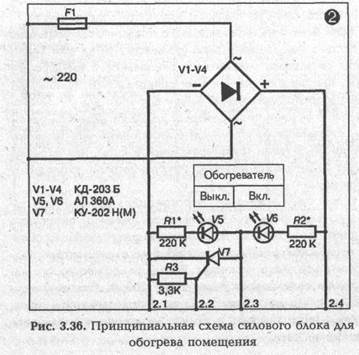
Обогрев помещения производится от сети 220 В без трансформатора. Выпрямительный мост блока собран на силовых диодах V1—V4. Это могут быть любые диоды на ток 10 А с обратным напряжением не менее 300 В. Исполнительным элементом схемы является тиристор V7 типа КУ 202Н(М)
В качестве нагревательного элемента можно использовать любые бытовые электрорадиаторы, электроконвекторы, электроплитки закрытого типа мощностью 500—1 000 Вт. Если температура поверхности нагревателя будет больше 80—90 °С, то эту поверхность надо накрыть толстым металлическим листом. Указанной мощности нагревателя достаточно для поддержания необходимой для зимовки температуры 5 ± ГС в помещении объемом до 10—12 м3 с теплыми (деревянными, из пенобетона, керамзита и т.д.) стенами и хорошо утепленным потолком. В помещении большего объема мощность нагревателя с данным силовым блоком можно увеличить, но не более чем до 2 кВт. При этом силовые диоды V1—V4 и особенно тиристор V7 должны находиться на вертикально расположенных металлических радиаторах площадью 40—50 см2 каждый. Светодиоды V5, V6 используются для контроля работы схемы.
Если будет принято решение осуществлять автоматический обогрев только в помещении и не заниматься внут-риульевым подогревом, то в этом случае надо будет изготовить только силовой блок для обогрева помещения и блок терморегулирования с отдельным блоком питания для него.
Блок питания в этом случае изготавливается по аналогии с блоком питания для подогрева ульев. Но трансформатор Т1 может иметь мощность не более 5—10 Вт, вторичное напряжение 26—30 В. Диоды V1—V4 любые на ток не более 0,1 А. Силовой узел — тиристор V6, светодиоды V7, V8 и соответствующие сопротивления исключаются.
Также исключается диод V5. Выходное напряжение этого блока питания выставляется подбором типа стабилитрона или цепочки стабилитронов в цепи базы транзистора VT1 и может составлять от 27 до 30 В. Может использоваться и любой другой блок питания со стабилизатором на соответствующее напряжение и ток не менее 0,1 А.
Внимание!При сборке схемы терморегулирования для помещения надо иметь в виду, что все элементы силового блока будут находиться под опасным для жизни потенциалом 220 В. Под этим же потенциалом будут находиться и элементы блока терморегулирования. По этой причине все эти элементы должны быть надежно изолированы от корпусов этих блоков и недоступны для случайного прикосновения при работе. 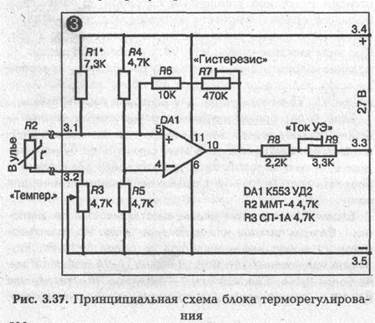
► Блок терморегулирования
Блок терморегулирования собран на микросхеме К553УД2. Термочувствительным элементом (термодатчиком) является термосопротивление R2 4,7 кОм типа ММТ-4, которое помещается в один из ульев над поверхностью подогревателя под низ рамок.
При настройке схемы потенциометром R7 «Гистерезис» выставляется ширина температурной петли гистерезиса в 1,5-2 °С. Делается это так: вращается ручка потенциометра R3 «Температура» в одну сторону до срабатывания схемы. На шкале «Температура» отмечается точка 1 срабатывания (включения) схемы. Затем потенциометр R3 вращается в противоположную сторону и отмечается точка 2 обратного срабатывания (выключения) схемы. Расстояние между точками 1 и 2 в °С и есть ширина температурного гистерезиса. Регулировкой R7 добиваются, чтобы разница этих температур была не больше 1,5—2 °С.
Потенциометром R1 «ток УЭ» выставляется ток управляющего электрода, при котором происходит надежное срабатывание (открытие тиристора). При этом надо проверить, будет ли при выставленном токе УЭ происходить закрытие тиристора. В моей схеме для тиристора КУ 202Л выставлен /уэ = 8 мА.
Подбором R1 выставляется значение выбранного температурного диапазона автоматического регулирования на шкалу «Температура», находящуюся на передней панели блока. При указанных на схеме значениях сопротивлений диапазон автоматического регулирования блока лежит в пределах от 0 до 50 "С. Градуирование шкалы «Температура» производится при помощи надежного термометра. Термодатчик R2 и термометр помещают в место с фиксируемой температурой на несколько минут. Необходимо иметь в виду, что тепловая инерционность термосопротивления в воздушной среде составляет не менее 120 секунд. Не меньшую инерционность имеет и термометр. Поэтому для надежного фиксирования температуры термосопротивление и термометр надо держать в среде с фиксируемой температурой не менее 8—10 минут. После этого вращением ручки потенциометра R3 добиваются выключения электроподогревателя ЕК и на шкале «Температура» делается соответствующая отметка. Температура, которую в это время показывает термометр, будет соответствовать сделанной на шкале отметке. Обращаю внимание на то, что шкала «Температура» должна градуироваться не на включение подогревателя, а на его выключение.Дело в том, что для одной и той же температуры, за счет тепловой петли гистерезиса, положение ручки на шкале «Температура» при включении и выключении подогревателя будет отличаться на 1,5—2 °С.
Указанное выше регулирование блока означает, что при работающей схеме терморегулирования в улье будет поддерживаться температура не выше той, которая будет выставлена на шкале.
В качестве фиксируемых температур можно пользоваться комнатной температурой, температурой над поверхностью хорошо прогретого электроподогревателя, температурой внутри холодильника, температурой тающего льда (0 °С) и др. В ходе эксплуатации аппаратуры градуирование шкалы «Температура» уточняется.
Для возможности выбора можно порекомендовать также оригинальную схему регулятора температур, которая приведена в № 9 журнала «Пчеловодство» за 2005 год на стр. 44.
Заканчивая рассмотрение вопроса, хочется дать совет тем, кто собирается изготовить комплект аппаратуры для терморегулирования: прежде чем начать изготовление, обязательно прочитайте ту часть приложения 3, в которой говорится о способах подключения нагрузок.
И последнее. Обращаю внимание на то, что представленная схема терморегулирования может с успехом использоваться в быту и для других целей: регулирования температуры в инкубаторах, аквариумах и т.д.
ПРИЛОЖЕНИЯ
 Приложение 1. Живое дерево: Осень — зима — весна
Приложение 1. Живое дерево: Осень — зима — весна
Глядя на сбросившее листву дерево, многим из нас кажется, что жизнь в нем остановилась до следующей весны. Кое-кто ничуть не сомневается в этом, однако не все так просто и однозначно в тех процессах, которые происходят в таком, казалось бы, известном нам растении, как дерево...
Ученые многих стран, изучающие жизнедеятельность (физиологию) древесных растений, пока не пришли к единому мнению относительно того, как именно древесный организм реагирует на снижение температуры окружающей среды, какие процессы при этом и в какой последовательности происходят в капиллярах и живых клетках древесины, почему и при каких условиях происходит полное прекращение жизнедеятельности (вымерзание) растений.
При глубокой проработке этого вопроса возникает множество частных проблем. Вот только один пример — вода. Уж, кажется, что может быть проще!
У многих из нас еще со школьных лет прочно установился стереотип о том, что вода замерзает при температуре 0 "С. Однако оказывается, что самая обычная вода, находящаяся в стеклянной трубочке с микрокапилляром в 0,1 мм, замерзает при —20 °С (Чудинов Б.С., 1984). В то же время в древесном капилляре такого же диаметра она замерзает при более высокой температуре. Приемлемого объяснения этому факту пока никто не дал. Более того, имеется инфррмация о том, что при определенных условиях обычная вода может оставаться в жидком состоянии (так называемая переохлажденная вода) при температурах вплоть до -70 °С (Веретенников А:В., 1987).
Существует также и ряд других проблем, по которым пока нет однозначного понимания.
По всем этим причинам предлагаемая модель функционирования живого дерева лиственной породы в период времени осень — зима — весна не может претендовать на бесспорность. Однако, с учетом всей имеющейся информации по этому вопросу на сегодняшний день, эта модель представляется наиболее вероятной.
♦ ♦ ♦
Процесс подготовки дерева к зиме начинается задолго до ее наступления. Считается, что механизм предварительной подготовки дерева к зиме включается с началом сокращения светового дня, когда в дереве начинают приостанавливаться процессы роста. Затем в конце лета дерево вступает в состояние так называемого внутреннего покоя, при котором в древесном организме резко снижается интенсивность обмена веществ.
При дальнейшем устойчивом снижении температуры воздуха ниже +10 °С, чаще всего это происходит в августе, дерево переходит в состояние глубокого (органического) покоя, который для большинства древесных растений средней полосы длится до ноября — декабря (Сергеев Л.И., 1964). По другому источнику — до ноября — января (Лесная энциклопедия, 1986).
Осенью с наступлением холодов обменные процессы в дереве продолжают замедляться. Уменьшаются в связи с этим и возможности проводящей системы дерева па доставке влаги и питательных веществ к ветвям (особенно тонким) и листьям. Однако транспирация (испарение) влаги в атмосферу не прекращается, и для того чтобы уменьшить транспирацию и привести ее хоть в какое-то соответствие с возможностями по доставке влаги, дерево сбрасывает листву. Таким образом, дерево защищает себя от зимнего иссушения.
В процессе характерных для осени многократных перепадов температур происходит так называемое закаливание дерева. В ходе этого периода внутри живых клеток дерева начинают накапливаться защитные вещества в виде растворов Сахаров и некоторых других веществ. Эти защитные вещества создают в дереве такие условия, при которых сильно снижается вероятность образования льда внутри живых клеток. Но даже у наиболее морозостойких деревьев вся имеющаяся в тканях вода не может оставаться незамерзающей. Какая-то ее часть при длительном воздействии низких отрицательных температур все же будет замерзать. Самое главное, чтобы этот процесс происходил не в протопласте (оболочке) живой клетки, а в межклеточном пространстве, тогда протопласт будет подвергаться меньшей опасности получить механические повреждения.
В лабораторных условиях при постепенном ступенчатом понижении температуры удалось добиться чрезвычайно высокой морозоустойчивости дерева. Так, в одном из опытов И.И. Туманова ветка яблони, помещенная в переносную морозильную камеру с температурой —153 °С, весной цвела и даже плодоносила (по А.А. Веретенникову, 1987).
При дальнейшем понижении внешних температур отток воды в межклеточные пространства увеличивается; в этот момент времени там будет находиться ровно столько воды, сколько ее сможет кристаллизоваться при имеющейся в данный момент температуре.
Процесс закаливания дерева и накапливания воды в межклетниках происходит постепенно и, в зависимости от характера наступающей зимы, может занимать продолжительный период времени. В ходе этого периода сокодвижение в дереве еще происходит, хотя и в замедленном темпе.
С переходом внешних температур в область устойчивых отрицательных значений корнеобитаемый слой почвы постепенно промерзает, что приводит к уменьшению поглощаемой корнями влаги.
При дальнейшем охлаждении древесины имеющаяся в ней вода после небольшого переохлаждения начинает замерзать вначале в наиболее крупных капиллярах. Но свойства воды таковы, что процесс ее кристаллизации сопровождается выделением теплоты. Так, при замерзании одного грамма воды выделяется 335 Дж (80 кал) тепла. По этой причине температура древесины, в капиллярах которой начинает замерзать вода, повышается до 0 °С и стабилизируется на этом уровне. Объясняется данное явление тем, что в лед за единицу времени может превратиться ровно столько воды, сколько необходимо для того, чтобы выделяющаяся при этом теплота компенсировала отводящуюся за это же время теплоту при охлаждении древесины внешним воздухом. Температура в отдельных точках внутри древесины начнет опускаться ниже 0 °С лишь после того, как вся свободная вода около них замерзнет. •
Вода в капиллярах дерева (так называемая свободная вода) начинает замерзать только при охлаждении древесины до —2...5 °С (Чудинов Б.С., 1984). Другой источник утверждает, что в стволах древесных пород образование льда происходит при —2...4 "С, а при сухой атмосфере — при -8... 11 "С (Самыгин Г.А., 1974).
Однако с началом замерзания в стволе кристаллизуется только часть воды, а часть этой свободной (не связанной в клетках) воды в межклеточных пространствах остается в жидком состоянии и сокодвижение, хотя и очень замедленно, продолжается до еще более низких температур. Способность воды (пасоки) сохраняться в переохлажденном жидком состоянии в тканях древесины, наличие в них растворов органических и минеральных веществ, способствующих сохранению жидкого состояния воды (наподобие антифриза), своевременная «закалка» дерева все увеличивающимися перепадами температур предохраняют дерево от гибели даже при трескучих морозах.
Что же касается связанной в клетках влаги, то она начинает замерзать при ее охлаждении до —5...—16 °С (Самыгин Г.А., 1974), до -10...15 °С (Чудинов Б.С., 1984). Принимая во внимание значительную инерционность температурного поля в древесных стволах, особенно большого диаметра, это происходит, видимо, при понижении температуры в атмосфере до —15...—20 °С (Чудинов Б.С., 1984).
Количество кристаллизуемой в межклетниках воды постепенно увеличивается, и в случае наступления продолжительного периода низких отрицательных температур в декабре — январе дерево переходит в состояние вынужденного (экзогенного) покоя.
Это состояние характеризуется наличием в межклеточных пространствах заболони определенного количества замерзшей воды, пЪчти полным отсутствием сокодвижения в стволе и ветвях, продолжением кутикулярной (покровной) транспирации и, как следствие, постоянным дефицитом влаги в живых тканях дерева. Дыхание живых клеток, испытывающих дефицит влаги, продолжается постоянно, хотя и с резко уменьшившейся интенсивностью. Дерево снизило все свои жизнеобеспечивающие функции до минимально возможного уровня, который, однако, достаточен для возобновления этих функций с наступлением временного или постоянного потепления. Подсушенные живые клетки готовы в любой момент принять недостающую воду, находящуюся в межклеточных пространствах в замерзшем и переохлажденном состоянии, как только она нагреется и перейдет в жидкое агрегатное состояние.
Параллельно с описанными выше процессами в это же время в дереве происходит и другой процесс. Как показывают исследования (Крамер П., Козловский Т., 1983), с началом перехода дерева в состояние внутреннего покоя в конце лета начинает изменяться концентрация газов в стволе дерева: концентрация кислорода начинает повышаться, а углекислого газа — уменьшаться. К январю — февралю кислород уже может занимать более 15% общего объема ствола (сосна) и до 30% у тополя, в то время как углекислый газ и у сосны и у тополя занимает не более 5% объема ствола. Эти изменения концентраций газов вызываются изменениями обменных процессов, которые в свою очередь определяются понижением среднесуточных температур. При этом четко прослеживается обратная зависимость между значениями внешних температур и концентрациями О2 и прямая зависимость между температурами и концентрациями СО2. Наличие этого процесса означает, что объем ствола, например, тополя в зимнее время занят в таком соотношении: около 30% древесиной, около 30% — свободной и связанной водой в разных агрегатных состояниях и более чем на 40% — газом (О2+ СО2) (Крамер П., Козловский Т., 1983).
Газ в древесине заполняет те полости и пустоты, которые не заняты водой. Если бы газ находился в капиллярах вместе с водой и разрывал непрерывные водные потоки, то это сделало бы невозможным передвижение водных потоков, ибо обязательным условием для передвижения жидкости в капилляре является непрерывность и неразрывность водного потока.
Наличие в стволе зимнего дерева большого количества пустот, составляющих до 40% общего объема ствола и занятых газом, придает древесине хорошие теплоизолирующие свойства.
Что же касается возможности пополнения запасов воды мелкими ветвями в холодное время года (а именно они испытывают в это время наибольший дефицит влаги), то И.И. Туманов (1955) установил следующее. Срезанные побеги, хранившиеся зимой на открытом месте, содержат меньше воды, чем побеги, находившиеся в тех же условиях на растениях. Объясняется это тем, что даже в холодные месяцы происходит пополнение влаги в ветвях за счет притока воды из других частей дерева, где имеются запасы, влаги. Такое передвижение воды может быть и при слабых морозах, если в древесном организме остается еще достаточное количество воды в жидкой фазе. Пополнение водного дефицита возможно даже при значительных морозах. Оно наблюдается при солнечной погоде, когда надземные части могут заметно прогреваться по сравнению с окружающим воздухом. Таким путем тонкие веточки, имеющие большую поверхность испарения, периодически пополняют израсходованную воду за счет ее запасов в более массивных частях растений. Если длительное время из-за сильных морозов этого происходить не будет, то ветви будут повреждаться и в дальнейшем отмирать.
Основной, причиной повреждения древесины при замерзании растений считается обезвоживание цитоплазмы клетки, вызванное образованием льда в межклетниках. Все более и более оттягивая воду из цитоплазмы, лед увеличивается в объеме и начинает чисто механически повреждать поверхностные слои обезвоженной цитоплазмы.
Ю.З. Кулагин (1969) также указывает на то, что во время зимних оттепелей и слабых (менее 5—7 °С) морозов в побеги поступает вода из толстых скелетных ветвей и ствола, которая ликвидирует опасный водный дефицит, вызванный зимним иссушением.
Однако при наступлении продолжительного периода низких отрицательных температур (—15...—20 °С) дерево переходит в состояние вынужденного покоя, при котором сокодвижение в дереве будет в значительной мере замедленно, а в отдельные периоды очень низких температур и вовсе прекращаться.
Ранней весной с началом потепления при прогреве ствола до положительных температур возобновляется интенсивное сокодвижение, в первую очередь за счет запасов свободной влаги, хранящейся зимой в межклеточных пространствах. По мере прогрева почвы и корнеобитаемого слоя возобновляется поступление влаги от корней, включается механизм активного поглощения воды древесным организмом. Движущей силой активного поглощения является так называемое корневое давление. Корневая система при этом превращается в своеобразный насос, который гонит влагу по стволу. Механизм активного поглощения действует только ранней весной. В дальнейшем дерево переходит на пассивное поглощение влаги за счет кутикулярной (с поверхности дерева), а затем — и листовой транспира-ции, которая теперь будет основной движущей силой, вызывающей перемещение влаги в проводящей системе дерева.
Дата добавления: 2016-03-30; просмотров: 876;