Нервно-мышечная передача
Место, где мотонейрон вступает в контакт с мышечной клеткой, называется нервно-мышечным синапсом(рис. 9-1). Клеточные мембраны мотонейрона и мышечной клетки разделены узким промежутком (20 нм) — синаптической щелью.Когда потенциал действия деполяризует терминаль мотонейрона, ионы кальция поступают извне в цитоплазму нерва, что вызывает слияние синап-тических пузырьков с пресинаптической терминальной мембранойи высвобождение содержащегося в них ацетилхолина в синаптическую щель.

Рис. 9-1.Нервно-мышечный синапс
Молекулы ацетилхолина диффундируют через синаптическую щель и взаимодействуют с никоти-ночувствительными холинорецепторами(н-холи-норецепторами) специализированной части мембраны мышечной клетки — концевой пластинкой скелетной мышцы.
Каждый холинорецептор состоит из пяти белковых субъединиц, две из которых (α-субъедини-цы) одинаковы и способны связывать молекулы ацетилхолина (одна α-субъединица — одно место связывания). Если оба места связывания заняты двумя молекулами ацетилхолина, то конформация субъединиц изменяется, что приводит к кратковременному (на 1 мс) открыванию ионного канала, проходящего через толщу рецептора (рис. 9-2).
Через открытый канал начинают поступать катионы (натрий и кальций — извне внутрь клетки, калий — из клетки наружу), что вызывает появление потенциала концевой пластинки. Содержимое одного синаптического пузырька — квант ацетилхолина — вызывает миниатюрный потенциал концевой пластинки(1 квант — 10 000 молекул ацетилхолина). Если ацетилхолином занято достаточное количество рецепторов, то суммарный потенциал концевой пластинки становится достаточно мощным для того, чтобы деполяризовать постсинаптическую мембрану вокруг синапса. Натриевые каналы в этой части мембраны мышечной клетки открываются под воздействием разности потенциалов (в отличие от каналов в рецепторах концевой пластинки, которые открываются при взаимодействии с ацетилхолином). Возникающий потенциал действияраспространяется вдоль мембраны мышечной клетки и системы Т-трубочек, что вызывает открывание натриевых каналов и выброс ионов кальция из цистерн саркоплазматической сети. Высвобожденный кальций опосредует взаимодействие сократительных белков актина и миозина, что приводит к сокращению мышечного волокна. Количество высвобожденного ацетилхолина обычно значительно превосходит минимум, необходимый для развития потенциала действия. Некоторые заболевания нарушают процесс нервно-мышечной передачи: при миастеническом синдроме Итона-Ламберта высвобождается недостаточное количество ацетилхолина, при миастении (myasthenia gravis) снижено число холинорецепторов.
Субстратспецифический фермент ацетилхо-линэстераза быстро гидролизует ацетилхолин на уксусную кислоту и холин. Молекулы этого фермента (называемого также специфической,или истинной, холинэстеразой)фиксированы в концевой пластинке в непосредственной близости от холинорецепторов. В конечном счете ионные каналы закрываются, что приводит к реполяриза-ции концевой пластинки. Когда распространение потенциала действия прекращается, ионные каналы в мембране мышечного волокна тоже закрываются. Кальций поступает обратно в сарко-плазматическую сеть, и мышечное волокно расслабляется.
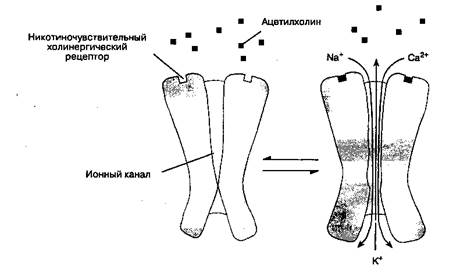
Рис.9-2. Связывание ацетилхолина с рецептором концевой пластинки скелетной мышцы приводит к открыванию канала и вызывает ионный ток
Дата добавления: 2016-03-30; просмотров: 751;