Рецепторный аппарат мышцы
Поперечнополосатая мышца – это не только конечное исполнительное звено в кортикомускулярном пути. Она является сложным механизмом, реализующим не только исполнительные команды, но и осуществляющим афферентную функцию в звене обратной связи: мышца-нервная система. Рецепторным аппаратом мышцы являются мышечные веретена и сухожильные рецепторы – сухожильные органы Гольджи. Мышечные веретена сигнализируют нервной системе о длине мышцы и о скорости изменения длины мышцы при ее сокращении; сухожильные органы Гольджи информируют о степени напряжения мышцы при сокращении. Мышечные веретена расположены продольно ходу мышечных экстрафузальных (вневеретенных) волокон в пределах тела мышцы (рис. 4).
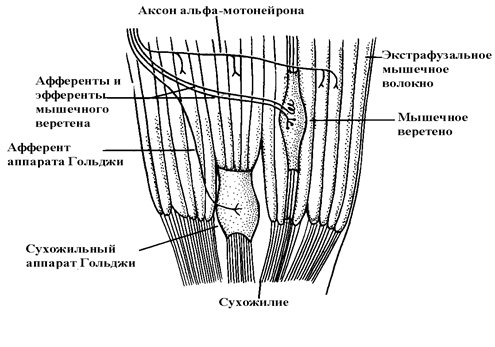
Рис. 4. Расположение мышечных веретен и сухожильных органов Гольджи
в скелетной мышце (Адаптировано по Houk et al., 1980).
Мышечное веретено представляет собой инкапсулированную структуру длиной от 4 до 10 мм, которая содержит три элемента: до 10-15 внутриверетенных (интрафузальных) волокон, рецепторные окончания и двигательный аксон. Интрафузальные волокна анатомически делятся на 2 типа: волокна с ядерной цепочкой и волокна с ядерной сумкой (рис. 5.).
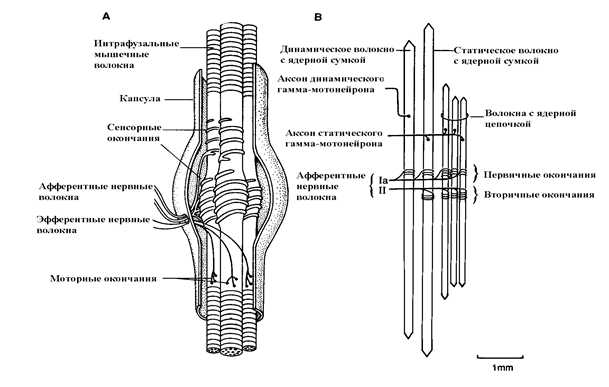
Рис. 5.Строение мышечного веретена. (Адаптировано по M.Hullinger, 1984 и I.A.Boyd, 1980).
А – мышечное веретено в разрезе,
В – взаимоотношения между интрафузальными мышечными и нервными афферентными и эфферентными волокнами.
Волокон с ядерной цепочкой в 2-3 раза больше, но они тоньше и короче, чем волокна с ядерной сумкой. Волокна с ядерной сумкой функционально делятся на динамические, то есть те, которые реагируют больше на скорость в изменении длины мышцы и на статические, реагирующие на изменение длины вне зависимости от скорости изменения длины мышцы. Интрафузальные волокна с ядерной цепочкой являются статическими. На экваториальной части всех 3-х типов интрафузальных волокон расположены Iа афференты (первичные нервные окончания) высокомиелинизированных волокон, которые информируют о длине мышцы и скорости ее изменения. Параэкваториально на волокнах с ядерной цепочкой и на статических волокнах с ядерной сумкой расположены II афференты (вторичные нервные окончания), менее миелинизированные, информирующие только о длине мышцы. Растяжение мышцы ведет к растяжению мышечных веретен, и за счет уменьшения диаметра волокон первичные и вторичные нервные окончания генерируют залпы активности, которые по отросткам униполярных чувствительных клеток моносинаптически активируют альфа-мотонейроны. Вторичные нервные окончания связаны с мотонейронами полисинаптически. Клинически в норме растяжение мышцы ведет к ее рефлекторному сокращению. Этот рефлекс был назван Шеррингтоном рефлексом растяжения (stretch reflex), или миотатическим рефлексом (myotatic reflex). Миотатический рефлекс имеет быстрый (фазический) компонент и более слабый и длительный тонический компонент. Фазический компонент обусловлен активацией первичными нервными окончаниями (Ia) альфа-больших мотонейронов, а тонический – активацией вторичными нервными окончаниями (II) альфа-малых мотонейронов. В основе всех сухожильных рефлексов лежит миотатический рефлекс. При денервации мышц миотатический рефлекс исчезает, а при (гамма) спастических парезах значительно повышается. Сопоставление данных Н-рефлекса и Т-ответа (сухожильного рефлекса) позволяет дифференцировать гамма-спастичность от альфа-спастичности и выявлять поражение мышечных рецепторов. Третьим структурным компонентом мышечного веретена являются моторные окончания гамма-мотонейронов на интрафузальных мышечных волокнах. В передних рогах спинного мозга наряду с альфа-малыми тоническими мотонейронами, которые иннервируют экстрафузальные тонические мышечные волокна (1 тип), и альфа-большими мотонейронами, которые иннервируют фазические мышечные волокна (2 тип), имеются гамма-мотонейроны - наиболее малые по размеру двигательные нейроны. Два вида гамма-мотонейронов – статические и динамические – иннервируют соответствующие интрафузальные волокна. Интрафузальные мышечные волокна имеют также поперечную исчерченность и по своим сократительным свойствам мало чем отличаются от экстрафузальных волокон. Активация гамма-мотонейронов приводит к сокращению концевых отделов интрафузальных мышечных веретен, в результате чего центральная часть растягивается и активируются мышечные рецепторы. Таким образом, через гамма-мотонейроны нервная система может регулировать чувствительность мышечных рецепторов. С учетом специфики иннервационной функции гамма-мотонейроны называют фузиомоторной системой, а альфа-мотонейроны - скелетомоторной системой. В условиях произвольного сокращения мышцы мышечное веретено становится расслабленным и может прекратить импульсацию, однако благодаря альфа-гамма коактивации этого не происходит, и даже в режиме сокращения мышцы интрафузальные мышечные волокна веретена приспосабливаются к изменившейся длине мышцы и продолжают импульсацию.
Сухожильные рецепторы Гольджи расположены на соединении сухожилия и мышцы, одним концом прикрепляясь к сухожилию, а другим вплетаясь в интерстиций мышцы. Рецепторы Гольджи представляют собой капсулу с коллагеновыми волокнами длиной до 1.5 мм. Один чувствительный аксон (Ib) входит в капсулу и разветвляется между коллагеновыми волокнами на множество ветвей, которые при сокращении мышцы и натяжении коллагеновых волокон сдавливаются и генерируют импульсы в спинной мозг. Импульсы от Ib афферентов в спинном мозге оказывают тормозное воздействие на альфа-мотонейроны через полисинаптические связи с участием тормозных нейронов.
Дата добавления: 2016-03-27; просмотров: 4487;