НЕРВНАЯ РЕГУЛЯЦИЯ ВЕГЕТАТИВНЫХ ФУНКЦИЙ
Со времени французского физиолога Биша — с начала XIX столетия — функции организма разделяют на аномальные, или соматические, и вегетативные. К анимальным, или соматическим, функциям относятся восприятие внешних раздражении и двигательные реакции, осуществляемые скелетной мускулатурой. Вегетативными функциями называют те, от которых зависит осуществление обмена веществ в целостном организме (пищеварение, кровообращение, дыхание, выделение и т. д.), а также рост и размножение. В соответствии с этим разделением функций различают соматическую и вегетативную нервную систему. Соматическая нервная система обеспечивает экстероцептивные сен-сорные и моторные функции организма. Вегетативная нервная система обеспечивает регуляцию деятельности внутренних органов, сосудов и потовых желез, а также трофическую иннервацию скелетной мускулатуры, рецепторов и самой нервной системы.
Вегетативная нервная система отличается от соматической локализацией своих ядер в ЦНС, очаговым выходом волокон из мозга, отсутствием сегментарности их распределения на периферии и малым диаметром волокон. Помимо этого, для вегетативной нервной системы характерно, что ее волокна, направляющиеся из мозга к внутренним органам, обязательно прерываются в периферических вегетативных ганглиях, образуя синапсы на нейронах, расположенных в этих ганглиях. Аксоны ганглионарных нейронов оказывают влияние на внутренние органы.
Ганглии являются не только образованиями, передающими импульсы из ЦНС на органы и ткани. Они представляют собой вынесенные на периферию рефлекторные центры, способные регулировать функции внутренних органов в известной мере независимо от ЦНС посредством периферических рефлексов, замыкающихся в этих ганглиях.
Вегетативная нервная система делится на симпатический и парасимпатический отделы. Они отличаются по локализации центров в мозге, по характеру влияний на внутренние органы, а также тем, что ганглии парасимпатического отдела расположены в самих иннервируемых ими органах (интрамурально), в то время как ганглии симпатического отдела локализованы в пограничном стволе (truncus sympaticus).
Выделяя регуляцию вегетативных функций, надо отметить, что в целостных реакциях организма сенсорные, моторные и вегетативные компоненты тесно связаны между собой.
Соматические компоненты реакций организма, осуществляемые скелетной мускулатурой, в отличие от вегетативных могут быть произвольно вызваны, усилены или заторможены; они находятся в течение всего хода реакции под контролем сознания.
Вегетативные же компоненты, как правило, произвольно не контролируются. На этом основании вегетативную нервную систему называют автономной, или непроизвольной. Однако представление об автономности вегетативной нервной системы является весьма условным.
Многочисленные опыты К. М. Быкова и сотрудников, показавшие возможность условнорефлекторной регуляции всех внутренних органов и всех вегетативных функций, позволяют считать, что кора больших полушарий регулирует деятельность всех органов, иннервированных вегетативной нервной системой, и координирует их деятельность в соответствии с текущими потребностями организма, в зависимости от характера его реакций при изменениях внешней и внутренней среды организма. Это обстоятельство свидетельствует о принципиальной возможности произвольного управления вегетативными функциями, что удается осуществить после специальной целенаправленной тренировки (например, по системе индийских йогов).
ОБЩИЙ ПЛАН СТРОЕНИЯ
И ОСНОВНЫЕ ФИЗИОЛОГИЧЕСКИЕ СВОЙСТВА
ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
Центры вегетативной нервной системы расположены в мозговом стволе и спинном мозге. 1. В среднем мозге находятся мезэнцефальные центры парасимпатического отдела вегетативной нервной системы; вегетативные волокна от них идут в составе глазодвига-тельного нерва. 2. В продолговатом мозге расположены бульбарные центры парасимпатического отдела нервной системы; эфферентные волокна от них проходят в составе лицевого, языкоглоточного и блуждающего нервов. 3. В грудных и поясничных сегментах спинного мозга (от I грудного до II — IV поясничного) находятся тораколюмбальные центры симпатического отдела вегетативной нервной системы: вегетативные волокна от них выходят через передние корешки спинномозговых сегментов вместе с отростками моторных нейронов. 4. В крестцовых сегментах спинного мозга находятся сакральные центры парасимпатического отдела вегетативной нервной системы, волокна от них идут в составе тазовых нервов.
Таким образом, центры вегетативной нервной системы расположены в четырех отделах ЦНС. Ядра, находящиеся в мезэнцефальном, бульбарном и сакральном отделах, образуют парасимпатическую часть вегетативной нервной системы, а находящиеся в тораколюмбальном отделе — ее симпатическую часть.
Все уровни вегетативной нервной системы подчинены высшим вегетативным центрам, расположенным в промежуточном мозге — в гипоталамусе и полосатом теле. Эти центры координируют функции многих органов и систем организма. Они в свою очередь подчинены коре больших полушарий, которая обеспечивает целостное реагирование организма, объединяя его соматические и вегетативные функции в единые акты поведения.
Симпатические нервные волокна имеют значительно более широкое распространение, чем парасимпатические. Симпатические нервы иннервируют фактически все органы и ткани организма; напротив, парасимпатические же нервы не иннервируют скелетную мускулатуру, ЦНС, большую часть кровеносных сосудов и матку.
Распространение волокон парасимпатического отдела вегетативной нервной системы представлено на рис. 104. Ко многим органам парасимпатические волокна проходят в составе блуждающих нервов, которые иннервируют бронхи, сердце, пищевод, желудок, печень, тонкий кишечник, поджелудочную железу, надпочечники, почки, селезенку, часть толстого отдела кишечника.
 Рис.104. Парасимпатический отдел вегетативной нервной системы (схема). Выделены: средний мозг, от которого берут начало парасимпатические волокна, приходящие в составе i.i.i,-зодвигательного нерва (III); продолговатый мозг, от которого отходят парасимпатические волокна в составе лицевого (VII), языкоглоточного (IX) и блуждающего (X) нервов; крестцовый отдел спинного мозга, от которого берет начало тазовый нерв.
Рис.104. Парасимпатический отдел вегетативной нервной системы (схема). Выделены: средний мозг, от которого берут начало парасимпатические волокна, приходящие в составе i.i.i,-зодвигательного нерва (III); продолговатый мозг, от которого отходят парасимпатические волокна в составе лицевого (VII), языкоглоточного (IX) и блуждающего (X) нервов; крестцовый отдел спинного мозга, от которого берет начало тазовый нерв.
|
 Рис.105. Симпатический отдел вегетативной нервной системы (схема). Сплошные линии — преганглионарные волокна, пунктирные — постганглионарные. Выделена тораколюм-бальная часть спинного мозга (от VII шейного до III поясничного сегмента), откуда берут начало преганглионарные симпатические волокна.
Рис.105. Симпатический отдел вегетативной нервной системы (схема). Сплошные линии — преганглионарные волокна, пунктирные — постганглионарные. Выделена тораколюм-бальная часть спинного мозга (от VII шейного до III поясничного сегмента), откуда берут начало преганглионарные симпатические волокна.
|
Распространение волокон симпатического отдела вегетативной нервной системы показано на рис. 105. Верхние сегменты симпатического отдела вегетативной нервной системы посылают свои волокна через верхний шейный симпатический узел к органам головы; следующие сегменты посылают их через нижележащие симпатические узлы к органам грудной полости и верхним конечностям; далее следует ряд грудных сегментов, посылающих волокна через солнечное сплетение и верхний брыжеечный узел к органам брюшной полости, и, наконец, от поясничных сегментов волокна направляются через нижний брыжеечный узел в основном к органам малого таза и нижним конечностям.
ДВУХНЕЙРОННАЯ СТРУКТУРА ЭФФЕРЕНТНЫХ СИМПАТИЧЕСКИХ И ПАРАСИМПАТИЧЕСКИХ ПУТЕЙ
Периферическая часть всех эфферентных симпатических и парасимпатических нервных путей построена из двух последовательно расположенных нейронов. Тело первого нейрона находится в ЦНС, его аксон направляется на периферию и оканчивается в том или ином нервном узле. Здесь находится тело второго нейрона, на котором аксон первого нейрона образует синаптические окончания. Аксон второго нейрона иннервирует соответствующий орган. Волокна первого нейрона называют преганглионарными, второго — постганглионарными.
Двухнейронная структура периферических эфферентных симпатических и парасимпатических путей является типичным признаком, отличающим их от соматических нервных волокон. На пути вегетативных нервов после выхода их из ЦНС, как правило, имеется только один перерыв нервного волокна, т. е. один синапс.
Из этого правила, однако, имеются некоторые исключения. Так, постганглионарные симпатические волокна, идущие к гладким мышцам желудочно-кишечного тракта, преимущественно оканчиваются не на мышечных волокнах, а на парасимпатических ганглиозных клетках, находящихся в стенке желудка и кишок. По-видимому, они снижают активность этих клеток и таким путем оказывают тормозящее влияние на гладкую мускулатуру. В данном случае, следовательно, имеется трехнейронная структура периферического пути. Исключением из отмеченного выше правила является также тот факт, что хромаффинные клетки мозгового слоя надпочечников иннервированы не постганглионарными, а преганглионарными симпатическими волокнами. Хромаффинные клетки, образующие под влиянием импульсов, поступающих к ним по симпатическим волокнам, адреналин, как бы заменяют постганглионарный нейрон, с которым они имеют общее происхождение. В данном случае имеется однонейронная структура эфферентного симпатического пути.
ГАНГЛИИ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
Ганглии симпатического отдела вегетативной нервной системы в зависимости от их локализации разделяют на вертебральные (иначе их называют паравертебральны-ми) и превертебральные. Вертебральные симпатические ганглии расположены по обе стороны позвоночника, образуя два пограничных ствола (их называют также симпатическими цепочками). Вертебральные ганглии связаны со спинным мозгом нервными волокнами, которые образуют белые соединительные ветви — rami communicantes albi. По ним к ганглиям идут преганглионарные волокна от нейронов, тела которых расположены в боковых рогах тораколюмбального отдела спинного мозга (рис. 106). Аксоны постгангли-онарных симпатических нейронов направляются от узлов к периферическим органам либо по самостоятельным нервным путям, либо в составе соматических нервов. В последнем случае они идут от узлов пограничных стволов к соматическим нервам в виде тонких серых соединительных веточек — rami communicantes grisei (серый их цвет обусловлен тем, что постганглионарные симпатические волокна не имеют миелиновых оболочек).
В ганглиях пограничного ствола прерывается большинство симпатических преган-глионарных нервных волокон; меньшая их часть проходит через пограничный ствол без перерыва и прерывается в превертебральных ганглиях.
Превертебральные ганглии распространяются на большем, чем ганглии пограничного ствола, расстоянии от позвоночника; вместе с тем они находятся в некотором отдалении и от иннервируемых ими органов. К числу превертебральных ганглиев относят солнечное сплетение, верхний и нижний брыжеечные узлы. В них прерываются симпатические преганглионарные волокна, прошедшие без перерыва узлы пограничного ствола.
Ганглии парасимпатического отдела вегетативной нервной системы расположены внутри органов или вблизи них (ресничный узел gangl. ciliare, ушной узел — gangl. oticum и некоторые другие). Аксон первого парасимпатического нейрона, находящегося в среднем мозге, продолговатом мозге или в сакральном отделе спинного мозга, доходит до иннервируемого органа не прерываясь. Второй парасимпатический нейрон расположен внутри этого органа или в непосредственной близости от него — в прилежащем узле. Внутриорганные волокна и ганглии образуют сплетения, богатые нервными клетками, расположенные в мышечных стенках многих внутренних органов, например сердца, бронхов, средней и нижней третей пищевода, желудка, кишечника, желчного пузыря, мочевого пузыря, а также в железах внешней и внутренней секреции.
Вегетативные ганглии играют важную роль в распределении и распространении проходящих через них нервных влияний. Число нервных клеток в ганглиях в несколько раз (в верхнем шейном симпатическом узле — в 32 раза, в ресничном узле — в 2 раза)
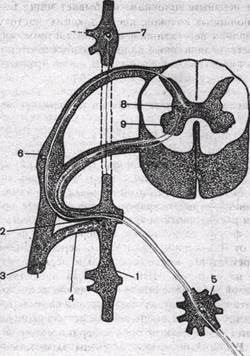 Рис.106. Связь симпатических ганглиев со спинным мозгом.
1 — вертебральный ганглий пограничного ствола;
2 — белая соединительная ветвь; 3 — смешанный нерв; 4 — серая соединительная ветвь; 5 — превертебральный ганглий; 6 — спинальный ганглий; 7—верхний шейный симпатический узел;
8 — симпатические нейроны; 9 — передний рог спинного мозга.
Рис.106. Связь симпатических ганглиев со спинным мозгом.
1 — вертебральный ганглий пограничного ствола;
2 — белая соединительная ветвь; 3 — смешанный нерв; 4 — серая соединительная ветвь; 5 — превертебральный ганглий; 6 — спинальный ганглий; 7—верхний шейный симпатический узел;
8 — симпатические нейроны; 9 — передний рог спинного мозга.
|
больше числа приходящих к ганглию пре-ганглионарных волокон. Каждое из этих волокон сильно ветвится и образует синапсы на многих клетках ганглия. Поэтому нервные импульсы, поступающие по преганглионарному волокну в ганглий, могут оказывать влияние на большое число ганглионарных нейронов и, следовательно, на еще большее число мышечных и железистых клеток иннервируемого органа. Таким образом достигается расширение зоны влияния преганглионарных волокон.
На каждом ганглионарном нейроне имеются синапсы, образованные многими преганглионарными волокнами. С этой конвергенцией связано явление пространственной суммации нервных импульсов. Если раздражать одно преганглионарное волокно стимулами подпороговой силы, то в постганглионарных волокнах не возникает потенциалов действия. Если же раздражать несколько преганглионарных волокон стимулами той же силы, какая применялась для раздражения одного волокна, то обнаруживается потенциал действия в постганглионарных волокнах в результате пространственной суммации постсинаптических возбуждающих потенциалов.
При раздражении преганглионарных нервных волокон отчетливо обнаруживаются также явления временной суммации нервных импульсов. Раздражение одиночным стимулом, как правило, не дает эффекта даже при большой силе раздражения. Раздражение ритмическими стимулами преганглионарных волокон вызывает возбуждение постганглионарных нейронов вследствие временной суммации постсинаптических возбуждающих потенциалов.
Одностороннее проведение нервных импульсов в межнейронных синапсах, перекрытие зон влияния отдельных входящих в узел преганглионарных волокон, наличие временной и пространственной суммации и окклюзии показывают, что нейроны и синапсы ганглиев вегетативной нервной системы обладают такими же свойствами, что нейроны и синапсы Ц.НС.
Наряду с этим при исследовании нейронов вегетативных ганглиев выявлен ряд существенных особенностей возникновения в них возбуждения. Одной из них является большая длительность синоптической задержки, составляющая от 1,5 до 30 мс (напомним, что синаптическая задержка в ЦНС составляет всего 0,3—0,5 с). Другая особенность нейронов вегетативных ганглиев состоит в большой длительности возбуждающего посте инаптического потенциала. Особенностью вегетативных нейронов является также резко выраженная в них следовая гиперполяризация, приводящая к возникновению депрессии вслед за волной возбуждения. С этими тремя особенностями возбуждения вегетативных нейронов связано то, что частота импульсов, которые они способны генерировать, относительно невелика— не превышает 10—15имп/с. Так, максимальный ритм импульсов, проходящих по сосудосуживающим нервным волокнам, не бывает чаще 6— 8 имп/с. Частый ритм возбуждений преганглионарных волокон, превышающих частоту естественных импульсов, возникающих в нейронах вегетативной нервной системы, частично блокируется в синапсах ганглиев и постганглионарные волокна возбуждаются в более редком ритме. Таким образом, обнаруживается трансформация ритма нервных импульсов. Частота стимуляции свыше 100 в секунду вызывает полную блокаду проведения через синапс.
Дата добавления: 2016-03-27; просмотров: 1910;