ВОЗБУЖДЕНИЕ В ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЕ
Хотя один и тот же химический медиатор, действуя на разные рецепторы постсинаптической мембраны в различных нервных клетках может вызывать как возбуждающие, так и тормозные процессы, в ЦНС позвоночных можно выделить синапсы, которые выполняют однозначную функцию—возбуждения (возбуждающие синапсы) (рис. 66). Так, центральные отростки первичных афферентных нейронов всегда оказывают возбуждающее действие на нейроны спинного мозга. Другим примером возбуждающего действия у позвоночных является мотонейрон, активирующий не только мышцы, но и вставочные клетки Реншоу спинного мозга.
В возбуждающих синапсах медиатор, высвобождаемый пресинаптическим окончанием, вызывает развитие локального процесса деполяризации, обозначаемого как возбуждающий постсинаптический потенциал (ВПСП). Указанное название подчеркивает тот факт, что ВПСП развивается в постсинаптической мембране.
В ЦНС млекопитающих ВПСП наиболее подробно изучены в спинальных мотонейронах, где имеется возможность избирательной активации однородных по составу афферентных волокон, которые образуют синапсы непосредственно на мотонейронах. Это позволяет изучать моносинаптические эффекты, не связанные с вовлечением в процесс возбуждения вставочных нейронов (рис. 67).
Амплитуда ВПСП зависит от исходного уровня мембранного потенциала. Смещение мембранного потенциала до величин, близких к нулю, обычно приводит к извращению (реверсии) знака ВПСП, т. е. суммарный постсинаптический ток в этих условиях течет в обратном направлении. Это означает, что активированная возбуждающим медиатором постсинаптическая мембрана становится проницаемой не только для ионов натрия, но и для некоторых других ионов, содержащихся внутри и снаружи клетки. Опыты с введением внутрь нейрона С1~ показали, что при этом амплитуда ВПСП не изменяется. По-видимому, возникновение ВПСП связано с одновременным увеличением проницаемости постсинаптической мембраны для Na+ и К+, а также, возможно, Са2+. Увеличение калиевой проницаемости приводит к уменьшению деполяризации, которая могла бы возникнуть за счет увеличения только натриевой или натриевой и кальциевой проницаемости.
Деполяризация нервной клетки в результате действия возбуждающего медиатора (ВПСП) может быть достигнута не только за счет увеличения проницаемости ее мембраны для Nа+ (или Са2+), но и за счет уменьшения проницаемости для К+.
Важным показателем эффективности синаптического возбуждения нервной клетки является способность возбуждающих синапсов вызывать возникновение потенциала действия. Необходимым условием для генерации потенциала действия является снижение трансмембранной разности потенциалов постсинаптической мембраны до определенного критического уровня.
Условия возникновения потенциала действия в нервной клетке под влиянием синаптического возбуждения в значительной степени обусловлены неодинаковой электрической возбудимостью различных участков мембраны и пространственным распределением различных возбуждающих синапсов. В большинстве центральных нейронов потенциал действия возникает в специальной низкопороговой области (обычно это зона аксонного холмика), откуда он распространяется по аксону и на мембрану соседних участков клетки. Указанный способ синаптического возбуждения нейрона очень важен для его интегративной функции, т. е. способности суммировать влияния, поступающие на нейрон по разным синаптическим путям.
ТОРМОЖЕНИЕ В ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЕ
Одним из фундаментальных свойств ЦНС является способность к торможению. Явление центрального торможения было открыто в 1863 г. И. М. Сеченовым, обнаружившим возникновение торможения спинальных центров лягушки при раздражении структур среднего мозга. В дальнейшем была выявлена возможность торможения спинальных реакций не только при раздражении надсегментарных образований, но и нервов противоположной стороны тела. Это открытие позволило подойти к установлению реципрокных отношений между процессами возбуждения и торможения в ЦНС. Реципрокный характер возбуждающих и тормозных влияний в спинном мозге показан учеником И. М. Сеченова Н. Е. Введенским и подробно проанализирован английским нейрофизиологом Ч. Шеррингтоном.
Важным шагом в выяснении природы центрального торможения оказалось выявление самостоятельного значения торможения для работы нервных центров. Торможение нельзя свести ни к утомлению нервных центров, ни к их перевозбуждению или католической депрессии нервных клеток. Торможение — самостоятельный нервный процесс, вызываемый возбуждением и проявляющийся в подавлении другого возбуждения. В отличие от процесса возбуждения, который может проявляться в двух основных формах — распространяющихся потенциалов действия и локальных потенциалов, торможение может развиваться только в форме локального процесса и поэтому всегда связано с существованием специфических тормозных синапсов (рис. 68).
Функция тормозных синапсов однозначна (они всегда вызывают только торможение), образующие их пресинаптические окончания относятся к аксонам так называемых тормозных нейронов, угнетающих активность всех нервных клеток, которые они иннервируют. Примером тормозных нейронов в спинном мозге являются вставочные нейроны Реншоу, в головном мозге — грушевидные нейроны (нейроны Пуркинье) коры мозжечка.
С помощью микроэлектродов установлено, что синаптическое торможение может вести к подавлению активности нейрона, имеющего тормозной синапс, вследствие изменения свойств постсинаптической мембраны нейрона (так называемое постсинаптическое торможение) или в результате уменьшения эффективности действия возбуждающих синапсов еще на пресинаптическом уровне (так называемое пресинаптическое торможение). Особенно широкое распространение в ЦНС имеет постсинаптическое торможение.
Постсинаптическое торможение. Медиатор, выделяемый пресинаптическими окончаниями тормозных синапсов, изменяет свойства постсинаптической мембраны таким образом, что способность нервной клетки генерировать процессы возбуждения (ВПСП или потенциал действия) подавляется. Поэтому данное явление принято обозначать как постсинаптическое торможение, а лежащее в его основе изменение в постсинаптической мембране—тормозной постсинаптический потенциал (ТПСП).
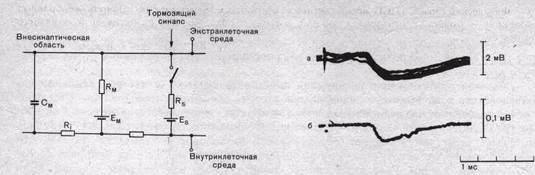
Рис. 68. Эквивалентная электрическая схема тормозного синаптического входа.
См—емкость мембраны; R^, R,—сопротивление мембраны; Rg—сопротивление синаптического контакта; Ем— электродвижущая сила мембраны, es — электродвижущая сила синаптического контакта.
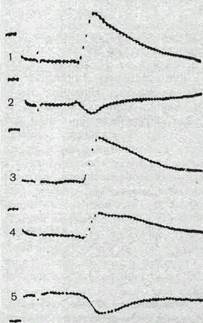
Рис.69. Тормозной постсинаптический потенциал (ТПСП) в мото нейронах спинного мозга кошки.
а — ТПСП, вызванный раздражением нерва;
б — ТПСП, вызванный раздражением вставочного нейрона.
Специфика тормозных синаптических эффектов была впервые наиболее подробно изучена на мотонейронах млекопитающих, а в дальнейшем — на многих нейронах мозга, включая нервные клетки мозгового ствола, гиппокампа и коры.
В мотонейронах спинного мозга возникновение ТПСП в ответ на раздражение афферентных волокон, идущих от мышц-антагонистов, обязательно связано с включением в тормозной процесс дополнительного звена — специального вставочного тормозного нейрона, аксональные окончания которого выделяют медиатор (вероятнее всего это глицин), вызывающий развитие ТПСП в постсинаптической мембране. Пример ТПСП в мотонейроне спинного мозга кошки показан на рис. 69. Как видно на этом рисунке, временное течение ТПСП.почти совпадает с временным течением ВПСП. И для ВПСП, и для ТПСП характерна более быстрая фаза нарастания и более длительная, убывающая по экспоненте фаза спада. ТПСП, возникающие при раздражении мышечных нервов, можно рассматривать как результат почти синхронного вовлечения совокупности тормозных нейронов. ТПСП, вызываемые прямым микроэлектродным раздражением одного тормозного нейрона, имеют сходные временные характеристики, но значительно меньшую величину.
Первоначально было сделано заключение, что торможение всегда развивается в результате гиперполяризации постсинаптической мембраны, так как тормозной медиатор увеличивает ее проницаемость для К+. В дальнейшем было установлено, что постсинаптическое торможение не обязательно сопровождается гиперполяризацией мембраны, так как более важное значение имеют лежащие в основе ТПСП сложные изменения ионной проводимости постсинаптической мембраны.
ТПСП обнаруживает очень высокую чувствительность к сдвигам мембранного потенциала, увеличиваясь при деполяризации и уменьшаясь при гиперполяризации. Когда последняя приводит к увеличению мембранного потенциала до 80 мВ, ТПСП превращается в деполяризационный ответ. Однако и в этом случае его тормозящее действие сохраняется.
Извращение ТПСП объясняется тем, что тормозной медиатор повышает проницаемость постсинаптической мембраны для С1-. В нормальных условиях концентрации С1- во внеклеточной среде превышает его содержание в нейроплазме. Во время развития ТПСП отрицательно заряженные ионы хлора устремляются внутрь клетки, увеличивая трансмембранную разность потенциалов. Когда концентрация Сl- в нейроплазме превышает его содержание в наружной среде, тормозной медиатор приводит к движению С1~ из клетки наружу, что приводит к ее деполяризации в результате потери отрицательных зарядов. Таким образом, тормозная постсинаптическая мембрана мотонейронов и других нейронов ЦНС действует как образование селективное к С1-, что, вероятно, обусловлено наличием положительных зарядов в стенках ионных каналов мембраны.
Физический смысл ТПСП всегда остается неизменным, он стремится сдвинуть мембранный потенциал в сторону, противоположную той, которая необходима для развития возбуждающего эффекта.
Учитывая природу тормозного процесса,.можно сделать вывод, что эффективность тормозных синапсов во многом зависит от их локализации на поверхности клетки. Тормозной эффект тем более значителен, чем ближе тормозной синапс расположен к месту генерации потенциала действия. Вследствие этого тормозные синапсы локализованы главным образом на теле нервных клеток вблизи от триггерной зоны аксонного холмика.
Поскольку функция тормозных синапсов заключается именно в подавлении или ограничении процессов возбуждения, развивающихся в постсинаптической мембране, важно рассмотреть особенности взаимодействия возбуждающих и тормозных постсинаптических эффектов.
В клетках ЦНС, получающих как возбуждающие, так и тормозные синаптические входы, их взаимодействие может быть рассмотрено на примере суммации ВПСП и ТПСП. Исследования, проведенные с помощью усреднения постсинаптических потенциалов на вычислительной машине, показали, что суммация ВПСП и ТПСП обычно имеет нелинейный характер (рис. 70).
Поэтому суммарная реакция нервной клетки на сочетанную активацию взаимодействующих входов значительно меньше алгебраической суммы обоих потенциалов. Наибольшая степень нелинейности наблюдается при совмещении начальных фаз ВПСП и ТПСП, т. е. в момент, когда лежащие в их основе изменения проводимости достигают максимума. Это полностью согласуется с тем, что эффект постсинаптического торможения обусловлен в первую очередь повышением проводимости постсинаптической мембраны.
Пресинаптическое торможение. Синаптическое торможение, приводящее к уменьшению эффективности возбуждающих синаптических влияний, может развиваться не только на уровне постсинаптической мембраны (как это было рассмотрено выше), но еще в пресинаптическом звене путем угнетения процесса высвобождения медиатора возбуждающими нервными окончаниями. В этом случае свойства постсинаптической мембраны не подвергаются каким бы то ни было изменениям.
Пресинаптическое торможение обнаружено в различных отделах ЦНС. Наиболее часто оно выявляется в структурах мозгового ствола и особенно в спинном мозге.
Так же как и постсинаптическое, Пресинаптическое торможение осуществляется посредством специальных тормозных вставочных нейронов.
|
Рис.70. Суммация возбудительного и тормозного постсинаптических потенциалов.
1 — ВПСП; 2—ТПСП'; 3—суммадия ВПСП и ТПСП'; 4 — суммация ВПСП и ТПСП"; 5— ТПСП".
Рис.71. Схема организации синапсов, участвующих в пресинаптическом торможении.
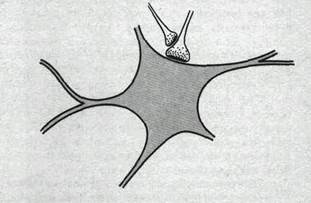
Структурной основой пресинаптического торможения являются аксоаксонные синапсы, образованные окончаниями аксонов тормозных вставочных нейронов и аксональными окончаниями возбуждающих нейронов. В этом случае окончание аксона тормозного нейрона является пресинаптическим по отношению к возбуждающему окончанию, которое в свою очередь будучи постсинаптическим по отношению к тормозному окончанию, является пресинаптическим по отношению к активируемой им нервной клетке (рис. 71). Импульсы в пресинаптическом тормозном аксоне высвобождают медиатор (в спинном мозге это вероятнее всего у-аминомасляная кислота), который вызывает деполяризацию возбуждающих окончаний за счет увеличения проницаемости их мембраны для С1-. Предполагается, что указанная деполяризация вызывает уменьшение амплитуды потенциала действия, приходящего в возбуждающее окончание, что в свою очередь уменьшает количество высвобождаемого им медиатора, вследствие чего амплитуда возбуждающего постсинаптического потенциала падает.
Другим механизмом пресинаптического торможения может быть уменьшение входящего внутрь потока Са2+, воздействующего на электросекреторную связь. И в этом случае пресинаптическое торможение приводит к уменьшению числа квантов медиатора, высвобождаемого возбуждающим пресинаптическим окончанием.
У млекопитающих продолжительность пресинаптического торможения значительно превосходит продолжительность постсинаптического торможения. Большая длительность пресинаптического торможения, по-видимому, обусловлена ритмической активностью тормозных вставочных нейронов.
Пресинаптическое торможение особенно эффективно при обработке информации, поступающей к нейрону по различным пресинаптическим путям. В этом случае возбуждение, поступающее по одному из синаптических входов, может быть избирательно уменьшено или даже полностью подавлено при отсутствии влияния на другие входы. Подобного результата нельзя достичь путем воздействия на проводимость постсинаптической мембраны, как это имеет место при постсинаптическом торможении, влияющем на всю нервную клетку.
ИНТЕГРАЦИЯ СИНАПТИЧЕСКИХ ВЛИЯНИЙ
Каждая клетка ЦНС имеет множество синаптических контактов с различными нейронами. Так, на одной клетке Пуркинье коры мозжечка насчитывают до 200 000 синапсов, число синапсов на мотонейронах млекопитающих составляет от 10000 до 20000.
Дивергенция. Способность нейрона устанавливать многочисленные синаптические связи с различными нервными клетками носит название дивергенции. Например, центральные окончания аксонов первичного афферентного нейрона образуют синапсы на многих мотонейронах-синергистах, на вставочных нейронах, осуществляющих торможение мотонейронов-антагонистов, и на клетках, дающих начало дорсальному спиноцеребеллярному восходящему тракту. Благодаря процессу дивергенции одна и та же нервная клетка может участвовать в различных нервных реакциях и контролировать большое число других нейронов, а каждый нейрон может обеспечивать широкое перераспределение импульсов, что приводит к иррадиации возбуждения.
Конвергенция. Схождение различных путей проведения нервных импульсов к одной и той же нервной клетке носит название конвергенции. Простейшим примером конвергенции служит факт получения каждым мотонейроном импульсов от совокупности первичных афферентных нейронов. На рис. 72 схематически показана конвергенция центральных окончаний первичных афферентных волокон на одном и том же мотонейроне и продемонстрирован конкретный пример развития ВПСП в мотонейроне 3 различными пресинаптическими элементами.
Если в приведенном выше примере конвергенция ограничивается эффектами, вызываемыми однотипными нейронами (все они относятся к сенсорным нейронам, проводя
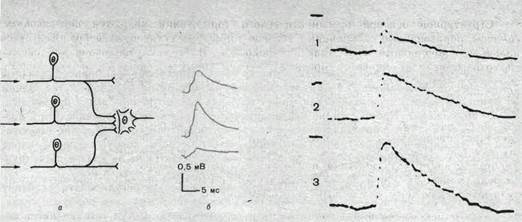
Рис.72. Схема конвергенции. Объяснение в тексте.
Рис.73. Линейная суммация ВПСП. Объяснение в тексте.
щим в спинной мозг импульсы от мышечных рецепторов растяжения), то значительная часть нервных клеток ЦНС имеет синапсы с нейронами различного типа, обеспечивающими конвергенцию влияний из разных источников. Например, к мотонейронам спинного мозга, кроме первичных афферентных волокон, конвергируют волокна различных нисходящих трактов, берущих начало в супраспинальных и собственно спинальных центрах, аксоны возбуждающих и тормозных вставочных нейронов. Поэтому мотонейроны рассматриваются как общий конечный путь многочисленных нервных структур, связанных с регуляцией моторной функции ЦНС. Принцип общего конечного пути был введен в физиологию нервной системы Ч. Шеррингтоном. Он показывает, каким образом одна и та же конечная реакция, проявляющаяся активацией определенной группы мотонейронов, может быть получена при раздражении различных нервных структур. Данный принцип имеет первостепенное значение для анализа рефлекторной деятельности (см. ниже) нервной системы.
Синаптическое взаимодействие. Конвергенция различных синаптических входов на одной нервной клетке обеспечивает возможность их взаимодействия. Так, при активации различных возбуждающих синапсов происходит пространственная суммация ВПСП. Пространственная суммация возбуждающих синаптических влияний имеет важное значение для возникновения импульсной активности в нервной клетке, так как деполяризации, создаваемой одним синаптическим входом, часто бывает недостаточно для достижения порогового уровня и генерации потенциала действия. Пространственная и временная суммация ВПСП способна приводить к длительной деполяризации постсинаптической мембраны, что обеспечивает возникновение ритмической импульсной активности нервной клетки.
Возникающие при активации различных синапсов ВПСП могут суммироваться линейно. При линейной суммации общая деполяризация равна арифметической сумме деполяризации, создаваемых каждым входом в отдельности. Возможна и нелинейная суммация, когда общая деполяризация нейрона меньше арифметической суммы деполяризации, создаваемых каждым возбуждающим синаптическим входом. Пример линейной суммации ВПСП, возникающих в одной и той же нервной клетке при раздражении разных пресинаптических путей, показан на рис. 73.
Линейный характер суммации наблюдается в том случае, когда взаимодействующие возбуждающие синапсы, конвергирующие на данном нейроне, расположены на таком расстоянии друг от друга, когда повышение проводимости постсинаптической мембраны, развивающееся под влиянием возбуждающего медиатора, не оказывает шунтирующего влияния на соседний вход. Наоборот, при достаточно близкой локализации взаимодействующих синаптических входов нервной клетки увеличение проводимости постсинаптической мембраны будет шунтировать и, следовательно, уменьшать деполяризацию, создаваемую соседним входом. Как отмечалось выше, суммация возбуждающих и тормозных постсинаптических потенциалов обычно развивается нелинейно. Чем выше степень нелинейности, тем сильнее выражен тормозной эффект.
Кроме взаимодействия непосредственно на постсинаптической мембране, различные синаптические влияния могут взаимодействовать еще и на пути к нервной клетке. Одним из примеров такого взаимодействия может служить рассмотренное выше пресинаптическое торможение. Кроме того, на пресинаптическом уровне может развиваться и процесс облегчения. Такое пресинаптическое или гетеросинаптическое облегчение заключается в увеличении эффективности одного синаптического возбуждающего входа в результате активации другого, когда облегчение развивается не на уровне постсинаптической мембраны нейрона, а вследствие взаимодействия на пути к нему.
Сами нервные клетки нередко обладают возможностью регулировать величину поступающих к ним сигналов. Такой механизм, получивший название обратной связи, заключается в том, что коллатерали аксонов нервной клетки могут устанавливать синаптические контакты со специальными вставочными нейронами, роль которых заключается в воздействии на нейроны или аксональные окончания путей, конвергирующих на нервной клетке, посылающей эти аксонные коллатерали. Так, например, возникновение импульса в мотонейроне млекопитающих не только активирует мышечные волокна, но и через коллатерали возбуждает специальные тормозные клетки Реншоу. Аксоны клеток Реншоу в свою очередь устанавливают синаптические связи с мотонейронами. Поэтому, чем сильнее импульсация мотонейрона, тем больше активируются клетки Реншоу и тем значительнее они тормозят мотонейроны, уменьшая частоту их импульсации (так называемое возвратное торможение).
Благодаря наличию обратных связей, степень возбуждения нейронов различных нервных центров может строго согласовываться как с интенсивностью приходящих к ним возбуждающих влияний, так и с интенсивностью импульсации на выходе нейронов и, следовательно, с интенсивностью развиваемого рабочего эффекта. Так, мотонейроны получают информацию о сокращениях мышцы от сухожильных и мышечных рецепторов. Эти импульсы, сигнализирующие о состоянии двигательного аппарата, позволяют корригировать активность этих мотонейронов. Афферентные импульсы, поступающие от сосудов, органов дыхания, пищеварения и выделения, постоянно корригируют деятельность нейронов, участвующих в поддержании уровня артериального давления и регуляции других вегетативных функций.
РЕФЛЕКТОРНАЯ ДЕЯТЕЛЬНОСТЬ ЦНС
Взаимодействие нервных клеток составляет основу целенаправленной деятельности нервной системы и прежде всего осуществления рефлекторных актов.
Как уже было сказано, принцип рефлекторной (отражательной) деятельности нервной системы был выдвинут еще в XVII в. французским философом и математиком Р. Декартом. Сам термин «рефлекс» был предложен в XVIII в. чешским физиологом Прохазкой. Весь последующий ход изучения деятельности нервной системы убедительно показал, что ее ответы на различные раздражения протекают по рефлекторному принципу. Рефлекторную природу психической деятельности обосновал И. М. Сеченов.
Рефлекторная дуга. Основой рефлекторного ответа является так называемая дуга рефлекса — комплекс специфически организованных нервных элементов, взаимодействие которых необходимо для осуществления рефлекторного акта. Рефлекторная дуга состоит из афферентной, центральной и эфферентных частей, связанных между собой с помощью синоптических соединений (рис. 74).
Афферентная часть представлена теми нервными элементами, которые формируют и проводят в центральном направлении нервные импульсы, необходимые для деятельности всей рефлекторной дуги. Поскольку возникновение афферентных импульсов связано с активацией специфических рецепторов, совокупность рецепторов, раздражение которых
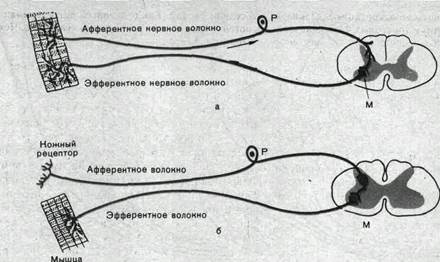
Рис.74. Схема двухнейронной (а) и трехнейронной (б) дуг спинномозгового рефлекса. Р—рецепторный нейрон спинального ганглия; М—мотонейрон.
вызывает определенный рефлекс, называют рецептивным полем рефлекса. Следует отметить, что раздражение одних и тех же рецепторов не всегда вызывает один и тот же тип рефлекторного ответа, а могут отмечаться различные рефлексы в зависимости от того, к каким центральным структурам импульсы поступают по первичным афферентным нейронам. Кроме того, в рецептивном поле одного рефлекса могут находиться и различные по функции рецепторы. Так, сгибательный рефлекс может вызываться раздражением тактильных рецепторов кожи или мышечных рецепторов.
Поступающие по афферентным путям нервные импульсы с помощью синаптических переключении активируют различные нейроны ЦНС. Часть афферентных импульсов, необходимых для возникновения рефлекторного ответа, переключается также на нейроны восходящих трактов и отражается в сознании. Однако многие рефлексы возникают и без участия сферы сознания, так как для их осуществления достаточно участие подкорковых зон ЦНС. Именно поэтому многие рефлекторные акты могут сохраняться даже после разрушения большей части ЦНС. Рефлекторная деятельность может осуществляться и одним изолированным сегментом спинного мозга, выделенным из организма и перфузируемым искусственным путем.
Моно- и полисинаптические рефлексы. Центральная часть рефлекса может включать несколько последовательных нейронов, соединенных синаптическими контактами. Тогда рефлекс носит название полисинаптического. В простейшем случае импульсы, поступающие в ЦНС по афферентному пути, переключаются непосредственно на эфферентный нейрон. Поскольку в пределах ЦНС рефлекторная дуга такого рефлекса имеет только один синапс (например, синапс между центральными окончаниями мышечных афферентов и мотонейронами), он носит название моносинаптического. Примером моносинаптического рефлекса является сухожильный рефлекс, или рефлекс растяжения.
В результате возникновения импульсов в эфферентных нейронах происходит активация эфферентной части рефлекса и его реализация. Рефлексы очень многообразны и их исполнительная часть включает различные органы и системы организма. В большинстве своем рефлексы служат для защиты организма и приспособления его к изменениям окружающей и внутренней среды. С их помощью адекватно координируются непроизвольные акты организма. Секреция желез, движения внутренних органов, реакция
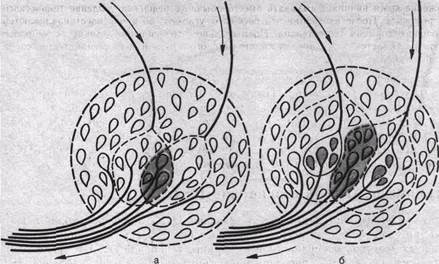
Рис.75. Окклюзия. Объяснение в тексте.
сердца и сосудов, скелетной мускулатуры тонко контролируются координированными рефлекторными актами.
Большинство детальных сведений о рефлекторных актах было получено благодаря изучению мышечных ответов, поскольку последние дают объективную и хорошо измеряемую оценку рефлекса. Примером двигательного рефлекса у человека является коленный рефлекс. Поскольку рефлекс не может быть вызван после того, как соответствующий центр разрушен или поврежден, наличие или отсутствие определенных рефлексов имеет важное диагностическое значение. Так, наличие коленного рефлекса свидетельствует о сохранности моторных центров поясничного сегмента спинного мозга. Рефлекторное сокращение зрачка в ответ на освещение показывает, что ядро III черепного нерва и соответствующие зрительные и эфферентные пути являются интактными.
Характер рефлекса в значительной степени зависит от интенсивности раздражения и числа активируемых рецепторов. Усиление раздражения приводит к расширению рецептивного поля рефлекса, в результате чего вовлекается большее число центральных нейронов. Указанное явление называется иррадиацией возбуждения. Процесс иррадиации в значительной степени зависит от того, что отдельные афферентные нейроны вызывают подпороговую деполяризацию центральных нейронов. При увеличении числа активированных афферентных нейронов в результате процессов пространственной суммации в большем числе центральных нейронов синаптическое возбуждение достигает порога и приводит к их импульсной активности.
Усиление раздражения вызывает также возрастание частоты импульсации в афферентных волокнах, что в свою очередь увеличивает ответы центральных нейронов вследствие временной суммации возбуждающих синаптических влияний.
Взаимодействие рефлексов. Различные рефлекторные реакции могут взаимодействовать между собой. Примером такого взаимодействия является феномен доминанты А. А. Ухтомского. Образование в ЦНС центра повышенной возбудимости приводит к тому, что раздражение самых различных рецептивных полей начинает вызывать рефлекторный ответ, характерный для деятельности этой доминантной области.
Доминантный очаг в ЦНС может возникать под влиянием разных факторов, например в результате гормональных воздействий. В частности, в период спаривания половые гормоны повышают возбудимость моторных центров шейного утолщения спинного мозга амфибий и любое раздражение кожи начинает вызывать вместо обычного рефлекса усиление тонического обнимательного рефлекса. После кастрации эти рефлексы угасают, но вновь восстанавливаются при введении полового гормона тестостерона. Показательно, что усиление активности нейронов моторных центров развивается даже при изоляции спинного мозга и добавлении тестостерона в перфузирующий раствор.
Взаимодействие рефлексов может проявляться как во взаимном облегчении (суммация), так и угнетении (окклюзия). Последнее явление выражается в том, что суммарный результат оказывается значительно меньшим, чем сумма взаимодействующих реакций. Согласно Ч. Шеррингтону (рис. 75), явление окклюзии объясняется перекрытием синоптических полей, образуемых афферентными частями взаимодействующих рефлексов. Поэтому при одновременном поступлении двух афферентных влияний ВПСП вызывается каждым из них отчасти в одних и тех же мотонейронах.
ОБЪЕДИНЕНИЕ НЕЙРОНОВ В НЕРВНЫЙ ЦЕНТР
В результате интегративной деятельности нейронов ЦНС, в основе которой лежат процессы суммации возбуждающих и тормозных постсинаптических потенциалов, возникающих в разных участках поверхностной мембраны, в нервной клетке происходит генерация импульсов или команд, регулирующих деятельность других нейронов или органов. В этом смысле работа индивидуального нейрона в какой-то степени подобна работе нервной системы в целом. Однако сложность и многообразие задач, выполняемых нервной системой, приводит к высокой специализации отдельных нейронов, и не только отдельных нейронов, но и нейронных объединений — нервных центров, деятельность которых связана с осуществлением различных функций. Функционирование ЦНС в целом осуществляется благодаря деятельности значительного числа таких центров, представляющих собой ансамбли нервных клеток, объединенных с помощью синаптических контактов и отличающихся огромным разнообразием и сложностью внутренних и внешних связей.
В соответствии с выполняемой функцией можно выделить различные чувствительные центры, центры вегетативных функций, двигательные центры, центры психических функций и т. д. Различные нервные центры характеризуются определенной топографией своего расположения в пределах ЦНС и более подробно будут рассмотрены в следующей главе.
Глава 6
ЧАСТНАЯ ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
В основе современных представлений о структуре и функции мозга лежит нейронная теория, рассматривающая ЦНС как совокупность синаптически связанных между собой нервных элементов — нейронов. Однако, несмотря на то что нервные клетки, из которых построен мозг, составляют единую особым образом организованную сеть, сложность ЦНС настолько велика, что для анализа ее функциональных свойств целесообразно разделение всего мозга на определенные области или отделы в соответствии с их макроструктурой, развитием, функцией и нейронной организацией. Такое подразделение в известной мере условно, так как нейроны, находящиеся в отдаленных друг от друга отделах мозга (например, гигантские пирамидные клетки двигательной коры и мотонейроны спинного мозга), бывают связаны между собой значительно более тесно, чем нейроны территориально близко расположенные, в пределах одной и той же области, но отно-
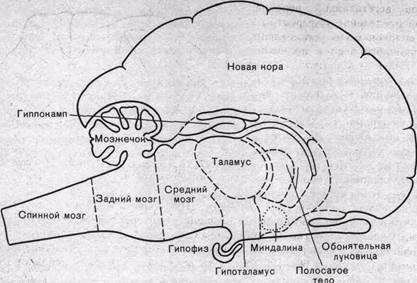
Рис.76. Основные отделы центральной нервной системы (схема).
сящиеся к различным физиологическим системам. Несмотря на это, разделение ЦНС на определенные отделы удобно как для нейрофизиологического анализа, так и для клинических целей.
Выяснение специфических особенностей различных нервных образований и закономерностей их взаимодействия относится к частной физиологии ЦНС.
Схематическое изображение ЦНС и входящих в ее состав отделов показано на рис. 76. На этой схеме в общем виде обозначены спинной мозг, задний мозг (включающий продолговатый мозг и варолиев мост или мост мозга), средний мозг, мозжечок, промежуточный мозг (таламус и гипоталамус) и передний мозг (включающий стриопаллидарную систему подкорковых ядер и кору больших полушарий).
СПИННОЙ МОЗГ
Спинной мозг — наиболее древний отдел ЦНС позвоночных, появившийся уже у наиболее примитивных представителей хордовых — ланцетника — еще до сформирования головного мозга. Поэтому он более, чем другие отделы ЦНС, сохранил черты древней организации, выражающиеся в метамерном строении.
Спинной мозг является самым каудальным отделом ЦНС. Он помещается в позвоночном канале и имеет неодинаковую длину у разных представителей позвоночных. У большинства рыб, хвостатых амфибий, рептилий и птиц он заполняет весь позвоночный канал, в то время как у бесхвостых амфибий и млекопитающих — только его часть. Поэтому в каудальном отделе позвоночного канала собираются корешки каудальных сегментов спинного мозга, образуя так называемый конский хвост.
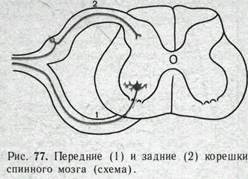
Спинной мозг характеризуется выраженным сегментарным строением, отражающим сегментарное строение тела позвоночных. От каждого спинномозгового сегмента отходят две пары вентральных и дорсальных корешков (рис. 77). Дорсальные корешки формируют афферентные входы спинного мозга. Они образованы центральными отростками волокон первичных афферентных нейронов, тела которых вынесены на периферию и находятся в спинномозговых ганглиях. Вентральные корешки образуют эфферентные выходы спинного мозга. В них проходят аксоны ос,- и у-мотонейронов, а также преганглио-
|
нарных нейронов вегетативной нервной системы. Такое распределение афферентных и эфферентных волокон было установлено еще в начале прошлого века и получило название закона Белла — Мажанди. После перерезки передних корешков на одной стороне наблюдается полное выключение двигательных реакций; но чувствительность этой стороны тела сохраняется. Перерезка задних корешков выключает чувствительность, но не приводит к утрате двигательных реакций мускулатуры.
Нейронная организация. Нейроны спинномозговых ганглиев относятся к простым
униполярным, или псевдоуниполярным, нейронам. Название «псевдоуниполярный» объясняется тем, что в эмбриональном периоде первичные афферентные нейроны происходят от биполярных клеток, отростки которых затем сливаются. Нейроны спинномозговых ганглиев можно подразделить на клетки малых и больших размеров. Тело крупных нейронов имеет диаметр порядка 60—120 мкм, в то время как у мелких нейронов он колеблется от 14 до 30 мкм.
Крупные нейроны дают начало толстым миелинизированным волокнам. От мелких начинаются как тонкие миелинизированные, так и немиелинизированные волокна. После бифуркации оба отростка направляются в противоположные направления: центральный входит в дорсальный корешок и в его составе — в спинной мозг, периферический — в различные соматические и висцеральные нервы, подходящие к рецепторным образованиям кожи, мышц и внутренних органов.
Иногда центральные отростки первичных афферентных нейронов заходят в вентральный корешок. Это происходит при трифуркации аксона первичного афферентного нейрона, в результате которой его отростки проецируются в спинной мозг и через дорсальный и вентральный корешки.
Из всей популяции клеток дорсальных ганглиев примерно 60—70% относится к мелким нейронам. Это соответствует тому, что число немиелинизированных волокон в дорсальном корешке превышает число миелинизированных волокон.
Тела нейронов спинномозговых ганглиев не имеют дендритных отростков и не получают синоптических входов. Их возбуждение происходит в результате прихода потенциала действия по периферическому отростку, контактирующему с рецепторами.
Клетки спинномозговых ганглиев содержат высокие концентрации глутаминовой кислоты — одного из предполагаемых медиаторов. Их поверхностная мембрана содержит рецепторы, специфически чувствительные к у-аминомасляной кислоте, что совпадает с высокой чувствительностью к у-аминомасляной кислоте центральных окончаний первичных афферентных волокон. Малые нейроны ганглиев содержат вещество Р или соматостатин. Оба этих полипептида также являются вероятными медиаторами, высвобождаемыми окончаниями первичных афферентных волокон.
Каждая пара корешков соответствует одному из позвонков и покидает позвоночный канал через отверстие между ними. Поэтому сегменты спинного мозга принято обозначать по тому позвонку, возле которого из спинного мозга выходят соответствующие корешки. Спинной мозг принято также разделять на несколько отделов: шейный, грудной, поясничный и крестцовый, каждый из которых содержит по нескольку сегментов, В связи с развитием конечностей нейронный аппарат тех сегментов спинного мозга, которые их иннервируют, получил наибольшее развитие. Это нашло свое отражение в образовании шейного и поясничного утолщений. В области утолщений спинного мозга корешки содержат наибольшее количество волокон и имеют наибольшую толщину.
На поперечном срезе спинного мозга ясно выделяется центрально расположенное серое вещество, образованное скоплением нервных клеток, и окаймляющее его белое вещество, образованное нервными волокнами. В сером веществе различают вентральные и дорсальные рога, между которыми лежит промежуточная зона. Кроме того, в грудных сегментах различают также боковое выпячивание серого вещества—боковые рога.
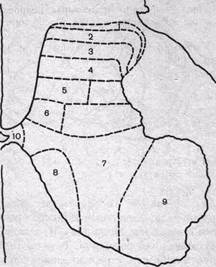 Рис. 78. Послойная топография серого вещества спинного мозга по Рекседу.
Цифрами обозначены слои нервных клеток.
Рис. 78. Послойная топография серого вещества спинного мозга по Рекседу.
Цифрами обозначены слои нервных клеток.
|
Все нейронные элементы спинного мозга могут быть подразделены на 4 основные группы: эфферентные нейроны, вставочные нейроны, нейроны восходящих трактов и интраспинальные волокна чувствительных афферентных нейронов. Моторные нейроны сосредоточены в передних рогах, где они образуют специфические ядра, все клетки которых посылают свои аксоны к определенной мышце. Каждое двигательное ядро обычно тянется на несколько сегментов. Поэтому и аксоны мотонейронов, иннервирующих одну и ту же мышцу, покидают спинной мозг в составе нескольких вентральных корешков.
Кроме моторных ядер, расположенных в вентральных рогах, выделяются большие скопления нервных клеток, в промежуточной зоне серого вещества. Это основное ядро вставочных нейронов спинного мозга. Аксоны вставочных нейронов распространяются как внутри сегмента, так и в ближайшие соседние сегменты.
Характерное скопление нервных клеток занимает также дорсальную часть дорсального рога. Эти клетки образуют густые переплетения, а указанная зона получила название желатинозной субстанции Роланда.
Наиболее точное и систематизированное представление о топографии нервных клеток серого вещества спинного мозга дает разделение его на последовательные слои, или пластины, в каждой из которых группируются главным образом однотипные нейроны.
Хотя послойная топография серого вещества была первоначально выявлена в спинном мозге кошки, она оказалась достаточно универсальной и вполне применима к спинному мозгу как других позвоночных, так и человека.
Согласно этим данным, все серое вещество можно разделить на 10 пластин (рис. 78). Самая первая дорсальная пластина содержит главным образом так называемые краевые нейроны. Их аксоны проецируются рострально, давая начало спиноталамическому тракту. На краевых нейронах оканчиваются волокна тракта Лиссауэра, который образован смесью первичных афферентных волокон и аксонами проприоспинальных нейронов.
Вторая и третья пластины образуют желатинозную субстанцию. Здесь локализуются два основных типа нейронов: более мелкие и относительно крупные нейроны. Хотя тела нейронов второй пластины имеют небольшой диаметр, их дендритные разветвления весьма многочисленны. Аксоны нейронов второй пластины проецируются на тракт Лиссауэра и собственный дорсолатеральный пучок спинного мозга, но многие остаются в пределах желатинозной субстанции. На клетках второй и третьей пластин оканчиваются волокна первичных афферентных нейронов, преимущественно кожной и болевой чувствительности.
Четвертая пластина занимает примерно центр дорсального рога. Дендриты нейронов IV слоя проникают в желатинозную субстанцию, а их аксоны проецируются в таламус и боковое цервикальное ядро. Синаптические входы они получают от нейронов желатинозной субстанции, а их аксоны проецируются в таламус и боковое цервикальное ядро. Синаптические входы они получают от нейронов желатинозной субстанции и первичных афферентных нейронов.
В целом нервные клетки первой — четвертой пластин захватывают всю вершину дорсального рога и образуют первичную сенсорную область спинного мозга. Сюда проецируются волокна большей части дорсально-корешковых афферентов от экстерорецепторов, включая кожную и болевую чувствительность. В этой же зоне локализованы нервные клетки, дающие начало нескольким восходящим трактам.
В пятой и шестой пластинах локализуются многочисленные типы вставочных нейронов, получающие синаптические входы от волокон заднего корешка и нисходящих путей, в особенности кортико-спинального и руброспинального тракта.
В седьмой и восьмой пластинах локализуются проприоспинальные вставочные нейроны, дающие начало длинным аксонам, достигающим нейронов отдаленных сегментов. Здесь заканчиваются афферентные волокна от проприорецепторов, волокна вестибулоспинального и ретикулоспинального трактов, аксоны проприоспинальных нейронов.
В девятой пластине располагаются тела а- и у-мотонейронов. Этой области достигают также пресинаптические окончания первичных афферентных волокон от мышечных рецепторов растяжения, окончания волокон нисходящих трактов, кортико-спинальных волокон, терминали аксонов возбуждающих и тормозящих вставочных нейронов.
Десятая пластина окружает спинномозговой канал и содержит наряду с нейронами значительное количество глиальных клеток и комиссуральных волокон.
СВОЙСТВА НЕРВНЫХ ЭЛЕМЕНТОВ СПИННОГО МОЗГА
Мотонейроны, расположенные в девятой пластине серого вещества, подразделяются на два основных типа: a- и у-мотонейроны. Альфа-мотонейроны иннервируют волокна скелетной мускулатуры (экстрафузальные волокна), обеспечивая мышечные сокращения. Гамма-мотонейроны иннервируют рецепторы растяжения (интрафузальные волокна). Благодаря сочетанной активации a- и у-мотонейронов рецепторы растяжения могут активироваться не только во время растяжения мышц, но и при их сокращении, что важно для обеспечения моторной координации.
Обнаружен также тип мотонейронов — (b-мотонейроны, концевые разветвления которых иннервируют параллельно экстрафузальные и интрафузальные волокна. Однако свойства этих мотонейронов изучены еще недостаточно.
Альфа-мотонейроны представляют собой крупные клетки с длинными дендритами. На соме и дендритах ос-мотонейронов размещается до 10 000—20 000 синапсов, большая часть которых образована окончаниями интраспинальных вставочных нейронов. Часть синапсов на а-мотонейронах представляет собой контакты с окончаниями первичных афферентных волокон, идущих от мышечных рецепторов растяжения, и с волокнами длинных нисходящих трактов: кортико-спинального, руброспинального, вестибулоспинального и ретикулоспинального.
Аксоны к-мотонейронов характеризуются высокой скоростью проведения — 70— 120 м/с, что согласуется с их значительным диаметром. Имеется четкая корреляция между размерами сомы мотонейронов, толщиной их миелинизированных аксонов и скоростью проведения по аксонам.
Деполяризация, возникающая в сх-мотонейронах при активации возбуждающих синаптических входов, вызывает ритмические разряды потенциалов действия. Частота импульсации пропорциональна степени деполяризации, однако обычно она не превышает 10—20 имп/с, что обусловлено интенсивной следовой гиперполяризацией, развивающейся после каждого потенциала действия.
Как и a-мотонейроны, у-мотонейроны расположены в девятой пластине, где они рассеяны среди а-мотонейронов. Гамма-мотонейроны представляют собой значительно более мелкие клетки. Диаметр их тела не превышает 30—40 мкм. В отличие от a-мотонейронов у-мотонейроны не имеют непосредственного синаптического контакта с первичными афферентными волокнами. Однако они моносинаптически активируются волокнами нисходящих трактов, что играет важную роль в а-, у-сопряжении (сочетанной активации).
В соответствии с незначительными размерами тела, аксоны у-мотонейронов также имеют небольшой диаметр. Поэтому скорость проведения по у-волокнам составляет всего 10—40 м/с. Следовая гиперполяризация в у-мотонейронах выражена незначительно и не ограничивает существенно частоты их импульсации. В этой связи у-мотонейроны могут разряжаться с частотой 300—500 имп/с.
| <== предыдущая лекция | | | следующая лекция ==> |
| МЕХАНИЗМЫ РЕГУЛЯЦИИ ФИЗИОЛОГИЧЕСКИХ ПРОЦЕССОВ | | | ПРОВОДЯЩИЕ ПУТИ СПИННОГО МОЗГА |
Дата добавления: 2016-03-27; просмотров: 8434;