МЕХАНИЗМЫ РЕГУЛЯЦИИ ФИЗИОЛОГИЧЕСКИХ ПРОЦЕССОВ
Человеческий организм представляет собой систему (грубее говоря — машину) — единственную по высочайшему саморегулированию. С этой точки зрения, метод изучения системы человека тот же, как и всякой другой системы: разложение на части, изучение значения каждой части, связи частей, соотношений с окружающей средой, и, в конце концов, понимание на основе всего этого ее общей работы и управление ею (И. П. Павлов). В этих словах выражено содержание понятия системного подхода.
Системный подход представляет собой методологию научного познания, в основе которого лежит рассмотрение объектов как систем. Этот подход ориентирует исследователя на раскрытие целостности объекта, на выявление многообразных типов связей в нем и на создание общего представления о системе. Объекты высокой степени сложности, к которым относится организм человека, представляют собой многоуровневую организацию, в которой системы более высокого уровня и сложности включают в себя системы более низкого уровня, образуя иерархию подсистем. Связи элементов в системе любого уровня осуществляются путем передачи информации. В организме животных и человека информация закодирована в определенной структуре биологических молекул, а также в определенном «рисунке» нервных импульсов (частота, набор в пачки, интервалы между пачками, определенное соотношение во времени импульсов и их пачек в различных нервных волокнах и т. д.).
С помощью передачи этой информации осуществляются процессы регуляции, т. е. управления физиологическими функциями, деятельностью клеток, тканей, органов, систем, поведением организма, осуществление взаимодействия организма и окружающей среды.
Главным регуляторным (управляющим) механизмом в организме высших животных и человека является нервная система. Основной механизм ее деятельности — рефлекс.
Рефлексом (от лат. reflecto — отражение) называют любую ответную реакцию организма, осуществляющуюся с участием центральной нервной системы. Морфологической основой таких реакций является рефлекторная дуга, включающая 5 звеньев:
1) рецептор—специализированный прибор, воспринимающий определенный вид воздействий внешней или внутренней среды; 2) афферентный (чувствительный) нейрон (или нейроны), проводящий сигнал, возникающий в рецепторе, в нервный центр; 3) вставочный нейрон (или нейроны), представляющий собой центральную часть рефлекторной дуги (или нервный центр) указанного рефлекса; 4) эфферентный (двигательный) нейрон, по аксону которого сигнал доходит до эффектора; 5) эффектор — поперечно-полосатая или гладкая мышца либо железа, осуществляющие соответствующую деятельность.
Любой эффектор, таким образом, связан элементами рефлекторной дуги с соответствующим рецептором и запускается в действие при раздражении данного рецептора. Ответная реакция организма возникает вследствие распространения по рефлекторной дуге возбуждения (сигнала), появляющегося при раздражении рецептора.
Понятие о рефлексе было введено в середине XVI века великим французским ученым Рене Декартом. Введение этого понятия сыграло важнейшую роль в развитии физиологии, позволило объяснить причину ответных реакций организма, изучить их механизм и показать, что в основе таких реакций лежит принцип детерминизма (т. е. Всеобщий как для неживой, так и для живой природы принцип причинно-следственных отношений). Тем самым был сделан важный шаг в развитии материалистических представлений о механизме реакций организма.
Со времен Декарта подобные реакции считались машинообразными, обеспечивающими автоматизированный ответ организма на раздражение рецептора. Однако подобные автоматизированные реакции имеют место лишь при возникновении элементарных простых рефлексов, которые могут осуществляться с участием ограниченных звеньев ЦНС.
Как правило, рефлекторные реакции организма являются гораздо более сложными и происходят при участии многих звеньев (этажей) ЦНС. Рефлексы при этом не сводятся к простым, однозначным ответным реакциям, а представляют собой звенья сложного процесса управления двигательными функциями (поведением) или деятельностью внутренних органов.
Функциональная структура таких процессов управления (регуляции) намного сложнее, нежели структура отдельных машинообразных рефлекторных ответов. Процессам управления независимо от того, где бы они не осуществлялись в организме животного или человека, производственном процессе, социальном обществе и т. д., присущи некоторые общие черты и закономерности.
Эти общие черты исследуются наукой, получившей название кибернетика. Кибернетика изучает общие черты и законы управления, осуществляемого на основе получения, хранения, передачи и переработки информации, независимо от физической природы объекта или системы, в которых осуществляются эти процессы. Кибернетическими системами могут быть автоматические регуляторы в технике, ЭВМ, организм человека и животных, биологическая популяция, человеческое общество.
Изучение законов кибернетики, понимание их смысла весьма важно для познания сущности процессов регуляции физиологических функций, для моделирования (математического или экспериментального) этих функций, для автоматического контроля за осуществлением этих функций, для вмешательства в физиологические процессы с целью их нормализации в случаях расстройств и заболеваний.
Изучение механизмов регуляции физиологических процессов раскрывает общность принципов кибернетики для всех указанных объектов, единство принципов автоматического регулирования в организме, в машине и производственном процессе.
Известно, что сами процессы управления и автоматического регулирования были использованы в технике гораздо раньше, чем они были открыты в организме, и до того, как были сформулированы законы кибернетики.
В машинах существуют «регуляторы, которые заменяют руку машиниста, приходя в целесообразную деятельность, как говорится сами собой, но в сущности под влиянием изменяющихся условий в ходе машины. Таков, например, предохранительный клапан в паровиках (паровых машинах) Уатта. По мере того, как напряжение пара в котле возрастает за известный предел, клапан сам собой увеличивает отверстие для выхода пара и наоборот. Таких приспособлений известно множество и все они носят название автоматических регуляторов. В животном теле, как в самодействующей машине, регуляторы, очевидно, могут быть только автоматическими, т. е. приводится в действие измененными условиями в состоянии или ходе машины (организма) и развивать деятельности, которыми эти неправильности устраняются» — писал И. М. Сеченов еще в 1897 году, предвосхищая положения кибернетики о механизмах саморегуляции в организме.
Таким образом, И. М. Сеченовым был сформулирован принцип отрицательной обратной связи, лежащий в основе процессов автоматического регулирования в машине и живом организме.
По этому принципу регулируются многие физиологические процессы. На значение этого факта впервые обратил внимание Клод Бернар (французский физиолог и патолог), обнаруживший значение постоянства внутренней среды для жизни организма. На примере регуляции уровня сахара в крови он показал, что любые отклонения этого уровня от нормы включают процессы, выравнивающие эти отклонения, что обеспечивает поддержание постоянства этой величины в организме. По этому же принципу регулируется постоянство температуры тела гомойотермных животных и другие параметры внутренней среды.
Немецкий ученый Карл Людвиг и русский физиолог Ф. И. Цион обнаружили подобный (работающий по принципу отрицательной обратной связи) механизм, регулирующий постоянство артериального давления в организме. Окончания чувствительного {депрессорного) нерва, локализованные в дуге аорты, при повышении давления крови в этом сосуде посылают усиленные сигналы в ЦНС. Эти сигналы вызывают рефлекторное замедление сердцебиения и расширение артериол, что приводит к падению артериального давления (т. е. к восстановлению его исходного уровня). Затем в организме было открыто большое количество подобных регуляторных механизмов. Значение в регуляции движений обратных связей, т. е. сигналов, поступающих из работающих мышц, подчеркнул И. М. Сеченов.
В ряде физиологических процессов был открыт механизм и положительной обратной связи, благодаря которой процесс, возникнув, усиливается и поддерживает сам себя.
Обратная связь — это связь на выходе системы. Она улавливает те или иные отклонения, уже возникшие в состоянии системы. Основанные на этом регуляторные механизмы работают по принципу «рассогласования». Деятельность их включается в тот момент, когда в состоянии системы уже наступают отклонения от заданной величины, т. е. когда возникает рассогласование между заданной (необходимой) и фактически возникшей величиной. Механизмы, работающие по этому принципу, широко распространены в организме. Общий принцип работы подобных механизмов представлен П. К. Анохиным в схеме «функциональной системы» (см. рис. 243). Подобная схема, однако, не является универсальной, т. к. в организме существуют регуляторные механизмы, работающие на основе иного принципа. Сигналом к их деятельности служит отклонение от заданной величины не на выходе, а на входе системы, т. е. действие на систему раздражителей, отличающихся от заданных параметров. В этом случае в основу регуляторных реакций положен иной принцип, т. е. работа регулятора «по возмущению». На входе системы имеются приборы, улавливающие величину поступающего сигнала, нарушающего состояние системы. Если эта величина превышает допустимую и может вызвать нежелательные отклонения в состоянии системы, то в таком случае возникают команды, обеспечивающие нейтрализацию действия этих сигналов и сохранение стабильного состояния системы. Здесь происходит не восстановление уже нарушенного состояния системы, а предупреждение возможности таких нарушений. (Оба эти принципа сохранения стабильности системы отличаются друг от друга, как, скажем, средства тушения уже возникшего пожара отличаются от средств и мер предупреждения пожаров.)
В любых физиологических регуляторных, защитных, компенсаторных реакциях имеет место взаимодействие обоих принципов и обоих механизмов регуляции, функционирующих как на выходе, так и на входе системы. Так, например, при воздействии на глаз струи пыльного воздуха, которая может вызвать засорение глаза, срабатывают (как почти и везде) оба механизма. Мигательный рефлекс, закрывая глаз, предупреждает попадание пыли (это механизм, работающий на входе системы «по возмущению»), а рефлекторное увеличение слезоотделения и промывание склеры и роговицы слезами удаляет уже попавшую пыль (механизм, работающий на выходе системы — «по рассогласованию»). В любой гомеостатической реакции можно наблюдать сочетание действия двух указанных механизмов, работающих на этих двух различных принципах.
Для любой регуляторной реакции необходимо получение информации о состоянии системы, о величине поступающих сигналов, о возникающих при этом сдвигах в ее состоянии. Необходим также аппарат сличения параметров этих сдвигов или параметров поступающих сигналов с величиной нормальных для данной системы параметров. Кроме того, необходим аппарат, формирующий команды, предотвращающие эти сдвиги. Действие этих команд осуществляется двумя путями: а) нормализацией уже возникших отклонений (механизмы, работающие «по рассогласованию»); б) предупреждением нежелательных эффектов входного (возмущающего) сигнала путем уменьшения силы сигнала, предотвращения его действия или снижения чувствительности системы к данному возмущающему воздействию (механизм, работающий «по возмущению»). Регуляторные реакции осуществляются в организме нервной системой.
Глава 5
ОБЩАЯ ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
Центральная нервная система координирует деятельность всех органов и систем, обеспечивает эффективное приспособление организма к изменениям окружающей среды, формирует целенаправленное поведение. Эти сложнейшие и жизненно важные задачи решаются с помощью нервных клеток (нейронов), специализированных на восприятии, обработке, хранении и передаче информации и объединенных в специфически организованные нейронные цепи и центры, составляющие различные функциональные системы мозга.
Объединение нервных клеток осуществляется с помощью синоптических соединений, важнейшей функцией которых является обеспечение перехода электрических сигналов с одного нейрона на другой.
Число нервных элементов, будучи очень ограниченным у примитивных организмов, в процессе эволюционного развития нервной системы достигает многих миллиардов у приматов и человека. При этом количество синаптических контактов между нейронами приближается к астрономической цифре — 1015—1016. Сложность организации ЦНС проявляется также в том, что структура и функция нейронов различных отделов мозга значительно варьируют. Тем не менее результаты исследования различных отделов мозга или клеток нервной системы животных, стоящих на разных уровнях эволюционного развития, позволяют выделить ряд общих закономерностей, определяющих течение основных нервных процессов: возбуждения и торможения в нейронах и синапсах ЦНС. Необходимым условием анализа деятельности мозга является выделение общих фундаментальных принципов, лежащих в основе функционирования нейронов и синапсов.
НЕЙРОННАЯ ТЕОРИЯ
В основе современного представления о структуре и функции ЦНС лежит нейронная теория, которая представляет собой частный случай клеточной теории. Однако, если клеточная теория была сформулирована еще в первой половине XIX столетия, то нейронная теория, рассматривающая мозг как результат функционального объединения отдельных клеточных элементов — нейронов, получила признание только на рубеже нынешнего века. До этого существовала тенденция рассматривать ЦНС как непрерывный синцитий, все элементы которого соединены прямой цитоплазматической связью.
Большую роль в признании нейронной теории сыграли исследования испанского нейрогистолога Р. Кахала и английского физиолога Ч. Шеррингтона. Окончательные доказательства полной структурной обособленности нервных клеток были получены с помощью электронного микроскопа, высокая разрешающая способность которого позволила установить, что каждая нервная клетка на всем своем протяжении окружена пограничной мембраной и что между мембранами разных нейронов имеются свободные пространства.
Нервная система построена из двух типов клеток: нервных и глиальных, причем число последних в 8—9 раз превышает число нервных. Однако именно нейроны обеспечивают все многообразие процессов, связанных с передачей и обработкой информации.
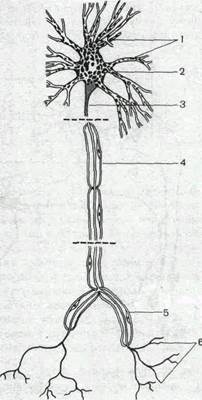 Рис.59. Нервная клетка (схема).
1 — дендриты; 2 — тело клетки; 3 — аксон-ный холмик, 4 — аксон; 5 — коллатераль аксона; 6—пресинаптические окончания аксона.
Рис.59. Нервная клетка (схема).
1 — дендриты; 2 — тело клетки; 3 — аксон-ный холмик, 4 — аксон; 5 — коллатераль аксона; 6—пресинаптические окончания аксона.
|
Основные структурно-функциональные элементы нервной клетки. В каждой нервной клетке можно выделить четыре основных элемента (рис. 59): тело, или сому, дендриты, аксон и пре-синаптическое окончание аксона. Каждый из этих элементов выполняет определенную функцию. Тело нейрона содержит различные внутриклеточные органеллы, необходимые для обеспечения жизнедеятельности всей клетки: ядро, рибосомы, эндо-плазматический ретикулум, пластинчатый комплекс (аппарат Гольджи), митохондрии. Здесь происходит основной синтез макромолекул, которые затем могут транспортироваться в дендриты и аксон. Мембрана тела большинства нейронов покрыта синапсами и, таким образом, играет важную роль в восприятии и интеграции сигналов, поступающих от других нейронов.
От тела клетки берут начало дендриты и аксон. В большинстве случаев дендриты сильно разветвляются. Вследствие этого их суммарная поверхность значительно превосходит поверхность тела клетки. Это создает условия для размещения на дендритах большого числа синапсов. Таким образом, именно дендритам принадлежит ведущая роль в восприятии нейроном информации. Мембрана дендритов, как и мембрана тела нейронов, содержит значительное число белковых молекул, выполняющих функцию химических рецепторов, обладающих специфической чувствительностью к определенным химическим веществам. Эти вещества участвуют в передаче сигналов с клетки на клетки и являются медиаторами синаптического возбуждения и торможения.
Основной функцией аксона является проведение нервного импульса — потенциала действия. Способность потенциала действия распространяться без ослабления обеспечивает эффективное проведение сигнала по всей длине аксона, которая у некоторых нервных клеток достигает многих десятков сантиметров. Таким образом, основная задача аксона — проводить сигналы на большие расстояния, связывая нервные клетки друг с другом и с исполнительными органами.
Окончание аксона специализировано на передаче сигнала на другие нейроны (или клетки исполнительных органов). Поэтому в нем содержатся специальные органеллы:
синоптические пузырьки, или везикулы, содержащие химические медиаторы. Мембрана пресинаптических окончаний аксона в отличие от самого аксона снабжена специфическими рецепторами, способными реагировать на различные медиаторы или нейромодуляторы. Благодаря указанному взаимодействию процесс выделения медиатора пресинаптическим окончанием может эффективно регулироваться другими нейронами. Кроме того, в отличие от остальной части аксона мембрана окончаний содержит значительное число кальциевых каналов, активация которых обеспечивает поступление внутрь окончания Ca2+.
Типы нейронов. Количество отростков, выходящих из тела нервной клетки, может значительно варьировать. В соответствии с этим различают уни-, би- и мультиполярные нейроны. Униполярные нейроны характерны главным образом для нервной системы беспозвоночных. В нервной системе позвоночных имеются преимущественно би- и
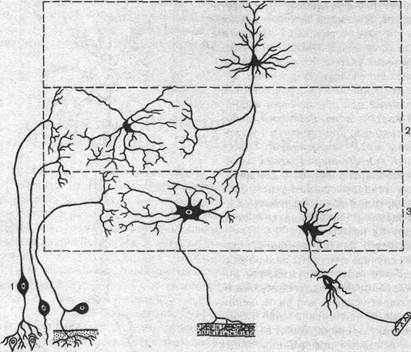
Рис.60. Типы нервных клеток. 1 — сенсорные нейроны; 2 — вставочные нейроны; 3 — эфферентные нейроны.
мультиполярные нейроны. Последние особенно характерны для ЦНС. Тела биполярных нейронов обычно расположены на периферии, но их центральные отростки вступают в ЦНС (рис. 60). Это так называемые первичные афферентные нейроны.
Различают 3 основных типа нейронов: афферентные, вставочные и эфферентные. Первичные афферентные нейроны воспринимают сигналы, возникающие в рецепторных образованиях органов чувств, и проводят их в ЦНС. Вступая в пределы ЦНС, окончания отростков первичных афферентных нейронов устанавливают синаптические контакты со вставочными, а иногда и непосредственно с эфферентными нейронами. Вставочные нейроны локализуются, как правило, в пределах ЦНС. Они обеспечивают связь между различными афферентными и эфферентными нейронами. Аксоны эфферентных нейронов, например мотонейронов, выходят за пределы ЦНС и иннервируют волокна скелетной мускулатуры. Многие нейроны, которые можно отнести к эфферентным, передают сигналы не прямо на периферию, а через посредство других нервных клеток. К таким эфферентным нейронам можно отнести нейроны различных отделов мозга, посылающие аксоны, идущие в составе длинных нисходящих трактов к спинному мозгу. Это пирамидные нейроны моторной зоны коры, руброспинальные, ретикулоспинальные и вестибулоспинальные нейроны, импульсы от которых поступают к двигательным клеткам спинальных моторных центров. Эфферентные нейроны вегетативной нервной системы расположены вне центральной нервной системы, в вегетативных ганглиях, находящихся на периферии. Их преганглионарные нейроны, локализованные в сером веществе мозгового ствола и спинного мозга, также относятся к эфферентным нейронам.
Кабельные свойства мембраны. Все многообразие электрических сигналов, генерируемых, перерабатываемых и посылаемых любой нервной клеткой, может быть сведено всего лишь к двум типам: локальным (градуальным) потенциалам и потенциалам дей
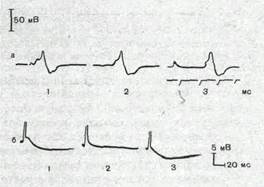 Рис.61. Потенциал действия, регистрируемый микроэлектродом, введенным в тело мотонейрона.
а — форма потенциала действия, вызываемого антидромно (1), синаптически (2) и прямым приложением тока через микроэлектрод (3); б— следовая гиперполяризация после потенциала действия мотонейрона котенка (1) и ее устранение после замены ионов кальция на ионы марганца (2) и восстановление в нормальном растворе (3).
Рис.61. Потенциал действия, регистрируемый микроэлектродом, введенным в тело мотонейрона.
а — форма потенциала действия, вызываемого антидромно (1), синаптически (2) и прямым приложением тока через микроэлектрод (3); б— следовая гиперполяризация после потенциала действия мотонейрона котенка (1) и ее устранение после замены ионов кальция на ионы марганца (2) и восстановление в нормальном растворе (3).
|
ствия (импульсным). Локальные потенциалы распространяются пассивно по кабельным структурам нейрона. Поэтому по мере удаления от места своего возникновения они затухают и могут служить только для проведения сигналов на сравнительно небольшие расстояния, например от тела или дендритов нервной клетки к области начального сегмента аксона, где обычно происходит процесс возникновения потенциала действия. Будучи значительно менее эффективным, чем потенциал действия, средством для передачи сигнала на расстояние, локальные потенциалы способны к суммации, и именно это свойство обеспечивает нейрону способность интегрировать все многочисленные поступающие к нему сигналы. Ввиду того что основным участком возникновения локальных потенциалов в нейронах ЦНС являются синапсы, которые расположены на мембране нейрона достаточно близко друг от друга, пространственное взаимодействие создаваемых синаптическими влияниями локальных процессов является достаточно эффективным.
Потенциал действия, служащий для проведения сигналов на большие расстояния, благодаря наличию регенеративного механизма распространяется без ослабления. Здесь же следует подчеркнуть, что в целой нервной клетке благодаря ее сложному геометрическому строению и неодинаковым свойствам мембраны в различных участках процесс возникновения и распространения потенциала действия отличается рядом особенностей.
Особенности возникновения и проведения потенциалов действия и локальных потенциалов. В большинстве нервных клеток порог возбудимости разных ее участков неодинаков. Он ниже всего в области аксонного холмика и начального сегмента аксона и выше в области сомы. Дендриты, как правило, имеют еще более высокий порог. Поэтому потенциал действия обычно возникает в области начального сегмента аксона и уже оттуда распространяется по аксону (ортодромно) и на тело клетки (антидромно). Если ввести в тело клетки микроэлектрод, позволяющий регистрировать потенциал действия, то можно видеть, что последний имеет характерную форму (рис. 61), демонстрирующую наличие двух основных компонентов. Первый компонент обусловлен активацией зоны начального сегмента и аксонного холмика, второй — тела и дендритов нейрона. Задержка между первым и вторым компонентами обусловлена тем, что более высокий порог возбудимости тела нейрона и значительное увеличение поверхности мембраны при переходе из аксонного холмика в тело нейрона затрудняют распространение потенциала действия на сомато-дендритическую мембрану.
После окончания потенциала действия во многих нейронах ЦНС наблюдается длительная следовая гиперполяризация. Она особенно хорошо выражена в мотонейронах спинного мозга.
Следовая гиперполяризация обусловлена тем, что соматическая мембрана в отличие от мембраны аксонов имеет значительное число кальциевых каналов. Деполяризация мембраны, развивающаяся во время потенциала действия, активирует кальциевые каналы соматической мембраны (П. Г. Костюк). Входящие внутрь клетки ионы кальция в свою очередь активируют калиевую проводимость мембраны.
Активация калиевой проводимости выражается в развитии следовой гиперполяризации, наблюдаемой после окончания потенциала действия. Если заменить ионы кальция в окружающей нейроны среде на ионы марганца, для чего необходимо осуществлять изоляцию и перфузию участка мозга, следовая гиперполяризация обратимо блокируется (рис. 61, б).
Следовая гиперполяризация играет важную роль в регуляции частоты потенциалов действия, генерируемых нервной клеткой. Способность нейрона отвечать ритмическими разрядами импульсов на длительную деполяризацию, создаваемую потоком импульсов, поступающих на его синапсы, представляет собой одну из важнейших характеристик его активности. В тех нейронах, где следовая гиперполяризация выражена значительно, частота импульсации не может быть очень высокой, так как ее верхние пределы ограничиваются фактически рефракторным периодом. Некоторые вставочные нейроны могут выдавать вспышки разрядов с частотой порядка 1000 в секунду. В мотонейронах спинного мозга длительность следовой гиперполяризации достигает 100—150 мс, что значительно увеличивает интервал между последующими потенциалами действия. Поэтому в обычных условиях частота ритмики мотонейронов не превышает 40—50 в секунду. Большинство двигательных актов осуществляется при еще более низкой частоте разрядов мотонейронов. Тонические мотонейроны имеют более длительную следовую гиперполяризацию и разряжаются с более редкой частотой, чем фазические мотонейроны, у которых следовая гиперполяризация короче.
МЕХАНИЗМЫ СВЯЗИ МЕЖДУ НЕЙРОНАМИ
Каждый многоклеточный организм, каждая ткань, состоящая из отдельных клеток, нуждается в механизмах, обеспечивающих межклеточные взаимодействия. Важное значение имеют процессы коммуникации клеток ЦНС. Главная задача их заключается в обработке и передаче информации, закодированной в виде электрических сигналов.
Хотя межнейронные взаимодействия могли бы осуществляться различными путями (например, с помощью влияния электрических полей, генерируемых близко расположенными нервными элементами, изменением ионного состава среды вследствие перераспределения ионов в результате предшествующей активности, выделением в окружающую среду различных продуктов обмена и т. д.), в основе деятельности мозга лежат в основном механизмы, обеспечивающие передачу электрических сигналов с нейрона на нейрон через межклеточные соединения — синапсы, специализированные на передаче этих сигналов. Являясь главным механизмом связи между нейронами, синапсы во многом обеспечивают все многообразие функций мозга.
Понятие синапс было введено в физиологию английским физиологом Ч. Шеррингтоном (1897) для обозначения функционального контакта между нейронами. Следует отметить, однако, что еще в 60-х годах прошлого столетия И. М. Сеченов подчеркивал, что вне межклеточной связи нельзя объяснить способы происхождения даже самого элементарного нервного процесса. Чем сложнее устроена нервная система и чем больше число составляющих мозг нервных элементов, тем более важное значение имеют синаптические контакты.
Структура и функция синапсов. Различные синаптические контакты отличаются друг от друга механизмом действия, локализацией на поверхности клетки, функциональной направленностью (возбуждающие или тормозящие), способностью к модуляции в результате предшествующей активности. Однако при всем многообразии синапсов существуют определенные общие свойства их структуры и функции. Поэтому прежде чем рассматривать специфические особенности синапсов различных отделов ЦНС, необходимо описать общие принципы их функционирования.
Синапс представляет собой сложное структурное образование, в котором следует различать пресинаптическое звено или пресинапс (чаще всего это концевое разветвление аксона) и постсинаптическое звено или постсинапс (чаще всего участок мембраны тела или дендрита другого нейрона). Кроме наиболее распространенных типов межнейронных контактов — аксосомагических и аксодендритических, существуют также аксоаксонные, дендродендритические, сомато-дендритические и дендросоматические синапсы.
Пресинаптическое окончание либо образует у постсинаптической клетки так называемые концевые бляшки, или бутоны, либо формирует по своему ходу многочисленные последовательные зоны контакта с различными участками постсинаптического нейрона (так называемые проходящие синапсы).
Механизм передачи через синапс долгое время оставался невыясненным, хотя было очевидно, что передача сигналов в синаптической области резко отличается от процесса проведения потенциала действия по аксону. В начале XX в. была четко сформулирована альтернатива: синаптическая передача осуществляется или электрическим, или химическим путем. Электрическая теория синаптической передачи в ЦНС пользовалась признанием до начала 50-х годов, хотя она и значительно сдала свои позиции после того, как химический механизм передачи был продемонстрирован в ряде периферических синапсов. Перфузия верхнего шейного симпатического ганглия (А. В. Кибяков), а также использование микроэлектродной.техники для внутриклеточной регистрации синаптических потенциалов нейронов ЦНС (Экклс) позволили сделать вывод о химической природе передачи в межнейрональных синапсах спинного мозга. Эти факты послужили основанием для вывода об универсальности химического механизма передачи во всех синапсах ЦНС.
Микроэлектродные исследования последних лет показали, однако, что в определенных межнейронных синапсах существует электрический механизм передачи. В настоящее время стало очевидным, что есть синапсы как с химическим, так и электрическим механизмом передачи. Более того, в некоторых синаптических структурах сочетание функционируют и электрический и химический механизмы передачи {смешанные синапсы, или синапсы двоякого действия). Синапсы с электрическим механизмом передачи чаще встречаются у животных с более примитивной нервной системой, хотя они и обнаружены в мозге млекопитающих, включая приматов. Их число уменьшается в процессе эмбрионального развития. Синапсы с химическим механизмом передачи составляют большую часть синаптического аппарата ЦНС высших животных и человека.
Структурные и функциональные особенности электрических, химических и смешанных синапсов. Для того чтобы потенциал действия, приходящий в Пресинаптическое окончание аксона, мог непосредственно возбудить постсинаптическую мембрану, т. е. вызвать в ней изменение мембранного потенциала, необходимо, чтобы значительная часть тока, текущего через пресинаптическую мембрану, могла входить в постсинаптическую клетку. Условием для такого вхождения тока является низкое сопротивление участка, связывающего обе клетки (они должны быть электрически связаны), и отсутствие шунтов, по которым пресинаптический ток мог бы ответвиться и не попасть на постсинаптическую мембрану.
Если синаптическая щель, разделяющая пре- и постсинаптическую мембраны, широкая (как это имеет место в химических синапсах, где она составляет в среднем 10—20 нм), подавляющая часть пресинаптического тока шунтируется низким сопротивлением щели и лишь примерно 0,0001 часть его попадает на постсинаптическую мембрану. Эта величина слишком мала, чтобы вызвать ощутимый сдвиг мембранного потенциала постсинаптического нейрона. Поэтому в синапсах с широкой синаптической щелью необходим другой механизм, способный изменить мембранный потенциал постсинаптической клетки. Таким механизмом является выделение пресинапсом особых химических веществ — медиаторов, которые, воздействуя на специфические рецепторы постсинаптической мембраны, способны изменять состояние ионных каналов постсинаптической мембраны. Изменение ионной проницаемости постсинаптической мембраны, в свою очередь, приводит к возникновению постсинаптического ионного тока, вызывающего падение напряжения на постсинаптической мембране — постсинаптический потенциал. Работа химического синапса схематически изображена на рис. 62, а. Таким образом, генератор постсинаптического. тока находится непосредственно в постсинапти
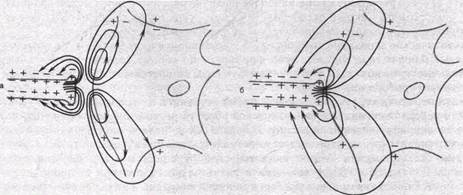
Рис.62. Схема передачи возбуждения в химическом (а) и электрическом синапсе (б). Стрелками показано распространение электрического тока через мембрану пресинаптического окончания и постсинаптическую мембрану на нейрон.
ческой мембране и запускается химическим медиатором, выделяемым пресинаптическим окончанием.
В электрических синапсах ширина синаптической щели составляет всего 2—4 им, что значительно меньше, чем в химических синапсах. Особенно важным является то, что в таких синапсах через синаптическую щель перекинуты мостики, образованные белковыми частицами. Они представляют собой своеобразные каналы шириной 1—2 нм, пронизывающие пре- и постсинаптическую мембраны синапса. Благодаря существованию таких каналов, размеры которых позволяют переходить из клетки в клетку неорганическим ионам и даже небольшим молекулам, электрическое сопротивление в области такого синапса (получившего название щелевого или высокопроницаемого контакта} оказывается очень низким. Это позволяет пресинаптическому току распространяться на постсинаптическую клетку без угасания. Поэтому механизм работы электрического синапса сходен в общих чертах с механизмом распространения волны деполяризации по нервному или мышечному волокну. Электрический ток течет от возбужденной области к невозбужденной и там вытекает наружу, вызывая ее деполяризацию (рис. 62, б). В электрическом синапсе потенциал действия достигает пресинаптического окончания и далее течет через межклеточные каналы, вызывая деполяризацию постсинаптической мембраны, т. е. генерируя возбуждающий постсинаптический потенциал (ВПСП). Важно подчеркнуть, что в электрическом синапсе генератор постсинаптического тока находится в пресинаптической мембране, где возникает активный процесс — потенциал действия. Из нее он пассивно (электротонически) распространяется на мембрану постсинаптической клетки. Поэтому синапсы с электрическим механизмом передачи часто обозначают как электротонические.
Структурная основа электрического синапса — высокопроницаемый щелевой контакт, обеспечивающий не только хорошую электрическую связь между нервными клетками, но и взаимный обмен различными органическими молекулами диаметром 1—2 нм. Более крупные молекулы, например белки, ДНК и РНК через межклеточные каналы не проходят. Однако и ограниченный обмен молекулами и ионами способен обеспечить определенную «метаболическую кооперацию» между нейронами, соединенными электрическими синапсами. Хотя электрические синапсы немногочисленны в ЦНС высших животных, они широко распространены в других возбудимых и невозбудимых тканях:
в сердечной мышце, гладкой мускулатуре внутренних органов в печени, эпителиальной и железистых тканях.
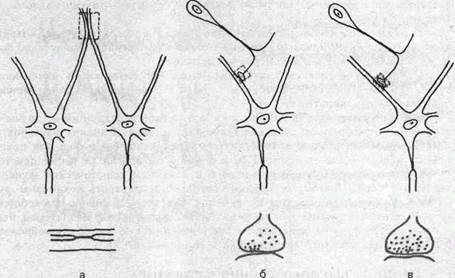
Рис.63. Схема чисто электрического синапса между дендритами мотонейронов лягушки (а), смешанного синапса между центральными окончаниями первичного афферентного нейрона и мотонейроном лягушки (б) и химического синапса между центральными окончаниями первичного афферентного нейрона и мотонейрона кошки (в).
В некоторых межнейронных синапсах электрическая и химическая передача осуществляются параллельно благодаря тому, что щель между пре- и постсинаптической мембранами имеет участки со структурой химического и электрического синапсов. Все 3 типа синапсов: электрический, химический и смешанный — схематически показаны на рис. 63. Обычно чисто электрические синапсы имеются между однотипными, близко расположенными нейронами, например между дендритами мотонейронов. Аксодендритические или аксосоматические синапсы, последовательно соединающие разные по функции и локализации нейро-ны, например первичные афферентные нейроны и мотонейроны, имеют химическую или смешанную природу.
Электрические и химические синапсы значительно отличаются друг от друга не только механизмом передачи, но и многими функциональными свойствами:
1. В синапсах с химическим механизмом передачи продолжительность сйнаптической задержки у теплокровных составляет 0,2—0,5 мс. В электрических синапсах синаптическая задержка, т. е. интервал между приходом импульса в пресинаптическое окончание и началом постсинаптического потенциала, отсутствует.
2. Химические синапсы отличаются односторонним проведением: медиатор, обеспечивающий передачу сигналов, содержится только в пресинаптическом звене. В электрических синапсах проведение чаще двустороннее, хотя геометрические особенности синапса делают проведение в одном направлении более эффективным. Кроме того, односторонность проведения в электрических синапсах может быть обеспечена полупроводниковыми свойствами мембраны.
3. Ввиду того что в химических синапсах возникновение постсинаптического потенциала обусловлено изменением ионной проницаемости постсинаптической мембраны, они эффективно обеспечивают как возбуждение, так. и торможение постсинаптического нейрона. В электрических синапсах активный процесс развивается в пресинаптическом звене, и поскольку нервный импульс всегда представляет собой волну деполяризации, электрические синапсы могут обеспечить передачу только одного процесса — возбуждения.
4. Химические синапсы значительно лучше, чем электрические, сохраняют следы предшествующей активности. Поэтому химическая передача значительно более подвержена модуляции под влиянием разных факторов.
5. Химические синапсы значительно более чувствительны к изменениям температуры, чем электрические, что имеет существенное значение для нервной системы пойкилотермных животных.
Поскольку химический механизм синаптической передачи имеет значительно более широкое распространение, чем электрический, детальный анализ факторов, определяющих передачу сигналов в химических синапсах, особенно важен для понимания различных аспектов деятельности ЦНС в норме и патологии (а также действия на мозг различных фармакологических веществ и токсинов, пластических функций нервной системы и т.д.). Поэтому необходимо детально рассмотреть механизмы функционирования синапсов с химическим механизмом передачи, а именно высвобождение медиатора пресинаптичеекими окончаниями, химическую природу медиаторов, молекулярную и ионную структуру их действия на постсинаптическую мембрану нейронов, лежащую в основе синаптического возбуждения и торможения.
ПРОЦЕСС ВЫСВОБОЖДЕНИЯ МЕДИАТОРА
Фактор, выполняющий медиаторную функцию, вырабатывается в теле нейрона и оттуда транспортируется в окончания его аксона, где в основном происходит его депонирование. Содержащийся в пресинаптических окончаниях медиатор должен выделиться в синоптическую щель, чтобы воздействовать на рецепторы постсинаптической мембраны, обеспечивая транссинаптическую передачу сигналов.
Еще до того, как были выяснены многие существенные особенности процесса высвобождения медиатора, было установлено, что пресинаптические окончания могут изменять состояние спонтанной секреторной активности. Выделяемые постоянно небольшие порции медиатора вызывают в постсинаптической клетке так называемые спонтанные миниатюрные пост синоптические потенциалы. Открытие спонтанного, т. е. не связанного с приходом нервного импульса, выделения медиатора помогло установить важнейшую особенность механизма его высвобождения — дискретный, квантовый характер. Дискретность процесса высвобождения выражается в том, что медиатор выходит из окончания не диффузно, не в виде отдельных молекул, а в форме многомолекулярных порций (или квантов), в каждой из которых содержится несколько тысяч молекул.
Постсинаптические эффекты, вызываемые в нервных клетках спонтанно выделяющимися квантами медиатора, наблюдаются особенно отчетливо в условиях, когда импульсная активность пресинаптических волокон искусственно подавлена, например, с помощью тетродотоксина — яда, избирательно блокирующего потенциалзависимые натриевые каналы мембраны, что устраняет возможность генерации потенциала действия. На рис. 64 показано, что после устранения импульсной активности спонтанные миниатюрные Постсинаптические потенциалы продолжают возникать через нерегулярные интервалы времени.
Приходящий в пресинаптическое окончание нервный импульс резко увеличивает высвобождение квантов медиатора. Возникающий в результате одновременного высвобождения многих квантов постсинаптический ответ, таким образом, представляет собой вызванный постсинаптический потенциал. Деполяризация пресинаптической мембраны является необходимым условием для высвобождения медиатора. Установлено, что такая деполяризация будет неэффективной, если в окружающей нейроны среде отсутствуют ионы кальция.
Действительно, если изолировать участок мозга и перфузировать его искусственным раствором, то при сохранении обычного ионного состава перфузирующей жидкости синаптическая передача в изолированном мозге не будет отличаться от передачи в условиях целого мозга и нор
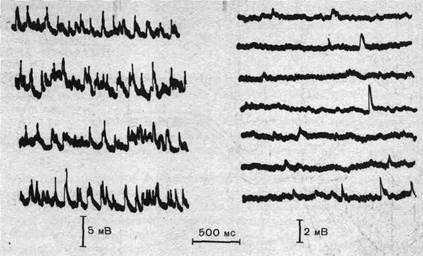
Рис.64. Спонтанные миниатюрные постсинаптические потенциалы, зарегистрированные при помощи внутриклеточного микроэлектрода в мотонейроне изолированного спинного мозга котенка.
а — при перфузии изотоническим раствором хлорида натрия; б — после выключения импульсной активности тетро до токсином.
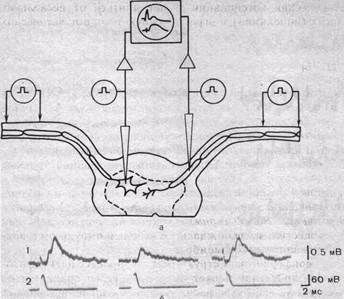
мального кровообращения. Удаление из перфузата ионов Са+ и особенно замена этих ионов на ионы Mg+ или Mn+, не влияя на спонтанное высвобождение квантов медиатора, прекращает высвобождение квантов медиатора нервными импульсами. Это особенно наглядно демонстрируют опыты на изолированном спинном мозге лягушки, поскольку здесь первичные афферентные волокна образуют смешанные синапсы со спинальными мотонейронами и возбуждающие постсинаптические потенциалы, возникающие в мотонейроне при раздражении одного такого афферентного волокна, содержат электрический и химический компоненты (рис. 65). Электрический компонент, отражающий возникновение потенциала действия в пресинаптическом окончании, не изменяется после удаления кальция из раствора. Напротив, медиаторный компонент полностью утрачивается. Таким образом, Са2+ является необходимым для процесса высвобождения медиатора нервным импульсом. При отсутствии Са2+ связь между деполяризацией пресинаптической мембраны и высвобождением медиатора (электросекреторная связь) нарушается. Роль кальция в этом процессе связана с тем, что деполяризация, создаваемая нервными импульсами, приводит к активации потенциалзависимых кальциевых каналов пресинаптической мембраны. Ионы Са, поступая внутрь пресинаптического окончания, обеспечивают выход квантов медиатора в синаптическую щель. Ионы Mg и Mn, блокируя потенциалзависимые кальциевые каналы мембраны, нарушают процесс высвобождения медиатора даже при наличии ионов Са в среде, окружающей клетки.
Ионы Са также участвуют и в спонтанном выбросе квантов медиатора, так как факторы, способствующие увеличению концентрации Са2+ внутри нервных окончаний, например некоторые метаболические ингибиторы, вызывают повышение частоты спонтанных миниатюрных потенциалов.
Дискретный, квантовый характер высвобождения медиатора нервным импульсом в синапсах ЦНС подтверждается результатами статистического анализа распределения амплитуд постсинаптических потенциалов, вызываемых раздражением одиночного пресинаптического волокна.
В разных синапсах ЦНС эффекты, вызываемые в постсинаптической мембране одиночным квантом медиатора, и средний квантовый состав, т. е. число квантов медиатора, освобождаемых в среднем нервным импульсом, значительно варьируют. Так, в синапсах между окончаниями первичных афферентных волокон и мотонейронами спинного мозга величина деполяризации, вызываемая одним квантом медиатора, обычно составляет 50—100 мкВ, а число квантов, высвобождаемых окончаниями одного волокна на данном мотонейроне, обычно не превышает 5—10, а часто бывает значительно меньше. В синапсах между некоторыми клетками коры мозжечка средний квантовый состав может достигать нескольких сотен.
| Рис.65. Схема опыта, позволяющего регистрировать постсинап-тические ответы в мотонейроне изолированного спинного мозга лягушки при раздражении одиночного пресинаптического волокна (а). Кривые (1, 2) иллюстрируют влияние удаления кальция на химический компонент такого элементарного постсинапти-ческого ответа. |
Электросекреторная связь. Электронно-микроскопические исследования показали, что пресинаптические окончания всегда содержат синаптические пузырьки или везикулы, каждая из которых содержит один квант медиатора. Действительно, имеются убедительные биохимические данные, что вещества, рассматриваемые в качестве химических медиаторов, содержатся в синаптических пузырьках. Более того, расчеты количества медиатора, содержащегося в одном пузырьке, и количество молекул медиатора, необходимых для создания постсинаптического эффекта, аналогичного действию одного кванта, совпадают. Таким образом, совокупность имеющихся данных свидетельствует о том, что как спонтанные миниатюрные постсинаптические потенциалы, так и постсинаптические потенциалы обусловлены выходом в синаптическую щель медиатора, содержащегося в синаптических пузырьках. Этот процесс (экзоцитоз) заключается в том, что пузырек, подойдя к внутренней поверхности мембраны пресинаптического окончания при наличии Са2+, сливается с пресинаптической мембраной. В результате происходит опорожнение пузырька в синаптическую щель. После спадения пузырька окружающая его мембрана включается в мембрану пресинаптического окончания, увеличивая его поверхность. В дальнейшем (в результате 'процесса эндоцитоза} небольшие участки пресинаптической' мембраны впячиваются внутрь, вновь образуя пузырьки, которые впоследствии снова способны включать медиатор и вступать в цикл его высвобождения.
Участие Са2+ в процессе высвобождения медиатора нервным импульсом определяет ряд важных специфических особенностей работы синапсов с химическим механизмом передачи. Такое характерное свойство химических синапсов, как синоптическая задержка, определяется главным образом временем, необходимым для вхождения Са 2+ внутрь пресинаптического окончания. Накопление Са 2+ внутри пресинаптического окончания в результате предшествующего поступления улучшает эффективность работы химического синапса. Если интервал между последовательным возникновением потенциалов действия в пресинапсе невелик, каждый последующий потенциал вызывает высвобождение большего числа квантов медиатора, что проявляется увеличением амплитуды постсинаптических потенциалов. Это явление временного облегчения или потенциации можно связать с накоплением Са2+ в пресинаптическом окончании. Такую же природу имеет и посттетаническая или постактивационная потенциация: увеличение числа квантов медиатора, высвобождаемых нервным импульсом, после предшествующего ритмического раздражения. Посттетаническая потенциация может длиться от нескольких минут до многих часов (в синапсах гиппокампа) и играть важную роль в пластических изменениях функции синапсов.
ХИМИЧЕСКИЕ МЕДИАТОРЫ
В ЦНС медиаторную функцию выполняет не одно, а большая группа разнородных химических веществ. Список вновь открываемых химических медиаторов неуклонно пополняется.
Чаще всего химическими медиаторами являются вещества с небольшой относительной молекулярной массой. Однако и высокомолекулярные соединения, такие, как полипептиды, также способны выполнять роль химических передатчиков в ряде центральных и периферических синапсов.
Основным критерием медиаторной функции веществ является его наличие в соответствующих пресинаптических окончаниях, способность высвобождаться под влиянием нервного импульса, а также идентичность молекулярных и ионных механизмов действия на постсинаптическую мембрану вещества, высвобождаемого нервным импульсом и прикладываемого искусственно к постсинаптической мембране.
В противоположность многим периферическим структурам, где процесс идентификации медиатора по указанным выше критериям может быть произведен сравнительно просто, ЦНС построена из негомогенных диффузно расположенных популяций нервных клеток и окончаний. Это вносит значительные трудности в обнаружение выделяемого медиатора, который, прежде чем появиться на поверхности мозга или в спинномозговой жидкости, должен диффундировать на большие расстояния. Помимо этого, в центральных структурах трудно добиться избирательной стимуляции определенной гомогенной группы нейронов или волокон, так же как трудно подводить предполагаемый медиатор к определенным нервным клеткам, не оказывая влияния на соседние нейроны. Именно поэтому природа химических медиаторов во многих синапсах ЦНС до сих пор окончательно не установлена. Тем не менее выявлен ряд веществ, играющих роль медиаторов синаптического возбуждения и торможения в ЦНС млекопитающих и человека.
К ним относятся: ацетилхолин; катехоламины: адреналин, норадреналин, дофамин; серотонин: 5-гидрокситриптамин; нейтральные аминокислоты: глутаминовая, аспарагиновая кислоты; кислые аминокислоты: глицин, гамма-аминомасляная кислота (ГАМК); полипептиды: вещество Р, энкефалин, соматостатин и др.; другие вещества: АТФ, гистамин, простагландины.
Согласно принципу Дейла, каждый нейрон во всех своих синаптических окончаниях выделяет один и тот же медиатор. Поэтому принято обозначать нейроны по типу медиатора, который выделяют их окончания. Нейроны, освобождающие ацетилхолин, называют холинергическими, серотонин — серотонинергическими и т. д. Этот же принцип может быть использован для обозначения различных химических синапсов. Иными словами, различают холинергические, серотонинергические и другие синапсы.
Ацетилхолин. Ацетилхолин является уксуснокислым эфиром холина, т. е. относится к простым эфирам. Он образуется при ацетилировании холина, причем этот процесс происходит при участии фермента ацетилхолинтрансферазы. Особенностью ацетилхолина как медиатора является быстрое его разрушение после высвобождения из пресинаптических окончаний с помощью фермента ацетилхолинэстеразы.
Ацетилхолин выполняет функцию медиатора в синапсах, образуемых возвратными коллатералями аксонов двигательных нейронов спинного мозга на вставочных клетках Реншоу, которые в свою очередь с помощью другого медиатора оказывают тормозящее воздействие на мотонейроны.
Этот пример является хорошей иллюстрацией принципа Дейла, так как известно, что периферические окончания моторных аксонов активируют волокна скелетной мускулатуры с помощью ацетилхолина. Возвратные коллатерали тех же аксонов в пределах ЦНС выделяют тот же медиатор.
Холинергическими являются и нейроны спинного мозга, иннервирующие хромаффинные клетки, а также преганглионарные нейроны, иннервирующие нервные клетки интрамуральных и экстрамуральных ганглиев. Полагают, что холинергические нейроны имеются в составе ретикулярной формации среднего мозга, мозжечка, базальных ганглиях и коре.
Катехоламины. Три родственных в химическом отношении вещества: дофамин, норадреналин и адреналин—.являются производными тирозина и выполняют медиаторную функцию не только в периферических, но и в центральных синапсах.
Дофаминергические нейроны находятся у млекопитающих главным образом в пределах среднего мозга, образуя так называемую нигростриальную систему. Особенно важную роль дофамин играет в полосатом теле, где обнаруживаются особенно большие количества этого медиатора. Кроме того, дофаминергические нейроны имеются в гипоталамусе.
Норадренергические нейроны содержатся также в составе среднего мозга, моста мозга и продолговатого мозга. Аксоны норадренергических нейронов образуют восходящие пути, направляющиеся в гипоталамус, таламус, лимбические отделы коры и в мозжечок. Нисходящие волокна норадренергических нейронов иннервируют нервные клетки спинного мозга.
Катехоламины оказывают как возбуждающее, так и тормозящее действие на нейроны ЦНС.
Серотонин. Подобно катехоламинам, серотонин относится к группе моноаминов, синтезируется из аминокислоты триптофана. У млекопитающих серотонинергические нейроны локализуются главным образом в стволе мозга. Они входят в состав дорсального и медиального ядер шва продолговатого мозга, моста и среднего мозга. Серотонинергические нейроны распространяют влияния на новую кору, гиппокамп, бледный шар, миндалину, подбугровую область, стволовые структуры, кору мозжечка, спинной мозг.
Серотонин играет важную роль в нисходящем контроле активности спинного мозга и в гипоталамическом контроле температуры тела. Нарушения серотонинового обмена, возникающие при действии ряда фармакологических препаратов, могут вызывать галлюцинации. Нарушения функции серотонинергических синапсов наблюдаются при шизофрении и других психических расстройствах. Серотонин может вызывать возбуждающее и тормозящее действие в зависимости от свойств рецепторов постсинаптической мембраны.
Нейтральные аминокислоты. Две основные дикарбоксильные кислоты L-глутамат и L-аспартат находятся в большом количестве в ЦНС и могут выполнять функцию медиаторов.
L-глутаминовая кислота представляет собой дикарбоновую аминокислоту, входящую в состав многих белков и пептидов. Она плохо проходит через гематоэнцефалический барьер и поэтому не поступает в мозг из крови, образуясь в самой нервной ткани (главным образом из глюкозы). В ЦНС млекопитающих глутамат обнаруживается в высоких концентрациях.. По-видимому, он является одним из самых распространенных медиаторов в центральных синапсах позвоночных животных. Полагают, что его функция связана главным образом с синаптической передачей возбуждения.
Глутамат исчезает из синаптической щели вследствие захвата его нервными и глиальными клетками и пресинаптическими окончаниями. Глутамат принимает участие в ряде важных метаболических процессов и входит в цикл синтеза у-аминомасляной кислоты. Сходное с ним действие оказывает на центральные нейроны аспартат.
Кислые аминокислоты. К этой группе аминокислот относятся ГАМК, и глицин.
ГАМК представляет собой продукт декарбоксилирования L-глутаминовой кислоты. Эта реакция катализируется декарбоксилазой глутаминовой кислоты. Отмечено значительное совпадение локализации этого фермента и ГАМК в пределах ЦНС. Другой фермент нервной ткани — трансаминаза — катализирует перенос аминогруппы ГАМК на сх-кетоглутаровую кислоту, в результате чего последняя превращается в семиальдегид янтарной кислоты.
| <== предыдущая лекция | | | следующая лекция ==> |
| ТРОФИЧЕСКАЯ ФУНКЦИЯ ДВИГАТЕЛЬНЫХ НЕРВНЫХ ВОЛОКОН И ИХ ОКОНЧАНИЙ | | | ВОЗБУЖДЕНИЕ В ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЕ |
Дата добавления: 2016-03-27; просмотров: 4250;