Главная функция промежуточных филаментов – поддерживающая и опорная.
Промежуточные филаменты в клетках различных типов различаются по своей химической природе и молекулярному весу. Выделяют 6 основных классов промежуточных филаментов
Цитокератины –промежуточные филаменты, характерные для клеток эпителия. Этот класс включает около 20 близких полипептидов (тонофиламентов). Кератиновые филаменты входят в состав десмосом и полудесмосом, участвуют в образовании рогового вещества в эпителии кожи и являются главным компонентом волос и ногтей.
Десмины – промежуточные филаменты мышечных тканей (за исключением миоцитов сосудов). Десмины играют важную роль в организации миофибрилл в мышечной ткани и обеспечении сократительной функции
Виментины – филаменты, характерные для различных клеток мезенхимного происхождения (фибробласты, макрофаги, остеобласты, эндотелий и гладкие миоциты сосудов).
Нейрофиламенты – промежуточные филаменты нейронов, которые играют важную роль в поддержании формы отростков нервных клеток.
Глиальные клетки содержат глиальный фибриллярный кислый белок и встречаются только в клетках нейроглии (астроциты, олигодендроциты).
Идентификация классов промежуточных филаментов (методами иммуноцитохимии с антителами к данному типу промежуточных филаментов) имеет большое значение в диагностике опухолей, и, следовательно, в прогнозе и выборе противоопухолевого лечения. Так, выявление различных форм кератинов свидетельствует о недифференцированных опухолях эпителиального происхождения, карциномах, аденокарциномах. Десмин является маркёром опухолей мышечного происхождения, а глиальный фибриллярный кислый белок – маркёр опухолей глиального происхождения.
ВКЛЮЧЕНИЯ
В отличие от органелл, включения цитоплазмы – непостоянные компоненты цитоплазмы, возникающие и исчезающие в зависимости от метаболического состояния клеток.
Включения подразделяются на трофические, секреторные, экскреторные и пигментные.
Трофические включения разделяются в зависимости от природы накапливаемого вещества на липидные, углеводные и белковые. Липидные включения – это капли нейтрального жира различного диаметра, которые накапливаются в цитоплазме и служат резервом энергетических субстратов, используемых клеткой. Из углеводных включений наиболее распространены гранулы гликогена (полимер глюкозы), эти включения также используются в качестве источника энергии. Примером белковых включений могут служить запасы белка вителлина в яйцеклетках животных. Они являются источником питания на ранних стадиях развития зародыша.
Секреторные включения имеют вид пузырьков, окруженные мембраной и содержащие биологически активные вещества, которые синтезируются в самой клетке, а затем выделяются (секретируются) во внешнюю среду. К таким включениям относятся секреторные гранулы, содержащие пищеварительные проферменты (зимогеновые гранулы), гормоны, медиаторы и др.
Экскреторные включения по своему строению сходны с секреторными, но в отличие от них, содержат вредные продукты метаболизма, подлежащие удалению из цитоплазмы клеток.
Пигментные включения представляют собой скопления эндогенных (синтезированных клеткой), или экзогенных (захваченных клеткой извне) окрашенных веществ - пигментов. Наиболее распространенными эндогенными пигментами являются гемоглобин, гемосидерин, билирубин, меланин, липофусцин; к экзогенным пигментам относят каротин, различные красители, пылевые частицы и др. Меланин – тёмно-коричневый пигмент, встречающийся в норме в коже, волосах, пигментной оболочке сетчатки в виде меланосом - гранул, окруженных мембраной. Липофусцин – гранулы жёлто-коричневого пигмента из продуктов лизосомного переваривания – накапливается в долгоживущих клетках (нейроны, кардиомиоциты), и поэтому его рассматривают как «пигмент старения».
ГИАЛОПЛАЗМА
Гиалоплазму называют также цитозолем, или клеточным матриксом. Гиалоплазма – сложная коллоидная система, которая может менять своё агрегатное состояние: переходить из более жидкого (золь) в более плотное (гель). Гиалоплазма состоит из гомогенного мелкозернистого вещества с низкой электронной плотностью, в которое погружены органеллы и включения. В составе гиалоплазмы – вода, белки (ферменты), нуклеиновые кислоты, полисахариды, липиды, а также неорганические вещества.
Функции гиалоплазмы:
· создание жидкой микросреды;
· метаболическая: метаболизм белков, жиров, углеводов.
III. ЯДРО. КЛЕТОЧНЫЙ ЦИКЛ. ДЕЛЕНИЕ КЛЕТКИ. КЛЕТОЧНЫЕ ПОПУЛЯЦИИ. ГИБЕЛЬ КЛЕТОК.
Ядро– важнейший компонент клетки, содержащий её генетический аппарат.
Функции ядра:
· хранение генетической информации (в молекулах ДНК, находящихся в хромосомах);
· реализация генетической информации, контролирующей различные процессы в клетке: транскрипция информационных, рибосомальных, транспортных РНК → синтетическая активность; апоптоз и т.д.);
· воспроизведение и передача генетической информации при делении клетки
В ядре неделящейся (интерфазной) клетки выявляются следующие 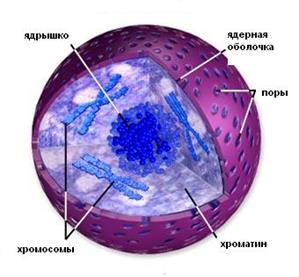 компоненты ядра:
компоненты ядра:
· ядерная оболочка (кариолемма);
· хроматин;
· ядрышко;
· кариоплазма.
Ядерная оболочка (кариолемма, нуклеолемма) на светооптическом уровне практически не определяется. Под электронном микроскопом обнаруживается, что она состоит из двух мембран – наружной и внутренней мембран, разделенных полостью шириной 15-40 нм – перинуклеарной цистерной.
Наружная мембранасоставляет единое целое с мембранами грЭПС: на её поверхности имеются рибосомы, а перинуклеарная цистерна сообщается с цистерной грЭПС
Внутренняя мембрана –гладкая, её интегральные белки связаны со слоем, состоящим из сети промежуточных филаментов (ламинов), - ламиной, или ядерной пластинкой. Ламина играет большую роль в поддержании формы ядра, укладке хроматина и структурной организации поровых комплексов.
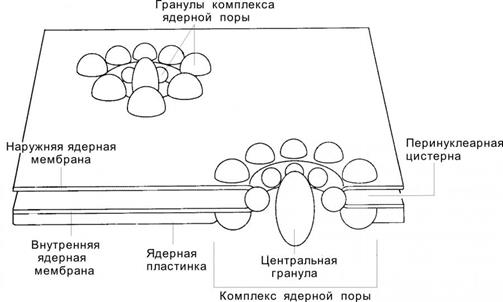
В определенных точках наружная и внутренняя мембрана смыкаются, образуя ядерные поры. Ядерная пора образована двумя параллельными кольцами диаметром 80 нм, содержащих по 8 белковых гранул, от которых к центру поры тянутся фибриллы, формирующие диафрагму толщиной около 5 нм. В середине диафрагмы лежит центральная гранула. Белковые гранулы ядерной поры структурно связаны с белками ламины. Совокупность компонентов, входящих в состав ядерной поры, называется комплексом ядерной поры.
Ядерная оболочка клетки содержит 2000-4000 поровых комплексов. Число поровых комплексов возрастает с увеличением функциональной активности: в клетках с высокой синтетической активностью ядерные поры занимают до 35% поверхности кариолеммы.
Комплекс ядерной поры обеспечивает избирательный транспорт веществ между цитоплазмой и ядром. По каналу, образованному поровым комплексом, движутся мелкие водорастворимые молекулы и ионы; активно переносятся в ядро белки, синтезируемые в цитоплазме (белки с маркировкой в виде с особой последовательности аминокислот – NLS, распознаваемой рецепторами NLS в комплексе поры); из ядра в цитоплазму переносятся субъединицы рибосом.
Хроматин в интерфазной (неделящейся) клетке соответствует хромосомам и состоит из комплекса ДНК и белка. Выраженность спирализации каждой из хромосом неодинакова по длине. Соответственно, различают два вида хроматина: эухроматин и гетерохроматин.
Эухроматин соответствует участкам хромосом, которые деспирализованы и открыты для транскрипции. Эти участки не окрашиваются и не видны в световой микроскоп.
Гетерохроматин соответствует конденсированным сегментам хромосом, что делает их недоступными для транскрипции. Гетерохроматин интенсивно окрашивается основными красителями, и в световом микроскопе имеет вид мелких гранул и глыбок.
По соотношению эу- и гетерохроматина в ядре можно оценить активность процессов транскрипции, и, следовательно, синтетической функции клетки. При её повышении это соотношение изменяется в пользу эухроматина, при снижении – нарастает содержание гетерохроматина. Соотношение эухроматин-гетерохроматин может, например, служить основой для дифференциальной диагностики доброкачественных и злокачественных опухолевых клеток.
При полном подавлении функции ядра в поврежденных и гибнущих клетках, оно уменьшается в размерах и содержит только гетерохроматин. Такое явление называется кариопикнозом.
Половой хроматин (тельце Барра) – скопление гетерохроматина, соответствующее одной из пары Х-хромосом, которая в интерфазе плотно скручена и неактивна.
Выявление полового хроматина используется как диагностический тест для определения генетического женского пола, что существенно при изучении генетических аномалий и, особенно, в спортивной медицине. Обычно анализируют эпителиальные клетки слизистой оболочки полости рта, где, как и в большинстве других клеток, половой хроматин выявляется как крупная глыбка гетерохроматина, лежащая рядом с ядерной оболочки. В нейтрофильных лейкоцитах крови половой хроматин имеет вид маленькой добавочной дольки ядра («барабанной палочки»).
Упаковка хроматина в ядре
В деконденсированном состоянии длина одной молекулы (двойной спирали) ДНК, образующей одну хромосому, составляет около 5 см, а общая длина молекул ДНК в ядре – более 2 м. Такие длинные нити ДНК компактно и упорядоченно упакованы в ядре диаметром всего 5-10 мкм.
Компактная упаковка молекул ДНК осуществляется благодаря связи ДНК со специальными основными белками – гистонами.
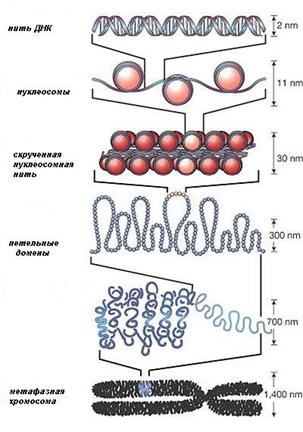 Начальный уровень упаковки хроматина – нуклеосома с
Начальный уровень упаковки хроматина – нуклеосома с
диаметром 11 нм.
· Нуклеосома состоит из блока, образованного комплексом из 8 молекул гистонов, на который намотана двойная нить ДНК (цепочка из 166 пар нуклеотидов).
· Нуклеосомы разделены короткими участками свободной ДНК (48 пар
оснований). Нуклеосомная нить имеет вид нитки с бусинами, где каждая бусина – нуклеосома.
· Второй уровень упаковки также обусловлен гистонами и приводит к скручиванию нуклеосомной нити (виток из 6 нуклеосом) с формированием хроматиновой фибриллы диаметром 30 нм.
· Хроматиновые фибриллы образуют петли диаметром 300 нм. При делении клетки в результате еще более компактной укладки и сверхспирализации ДНК появляются хромосомы (диаметр 700 нм), видимые под световым микроскопом.
Компактная упаковка ДНК в ядре обеспечивает упорядоченное расположение очень длинных молекул ДНК в небольшом объеме ядра, а также функциональный контроль активности генов.
Ядрышко выявляется в интерфазном ядре на светооптическом уровне как мелкая (~ 1 мкм в диаметре), плотная сферическая структура, интенсивно окрашивающееся основными красителями. Ядрышко образовано специализированными участками хромосом – ядрышковыми организаторами, на которых происходит синтез рибосомальной РНК, а также её сборка в предшественники рибосомальных субъединиц.
Компоненты ядрышка:
· Аморфный компонент, слабо окрашиваемый, представляет собой участки расположения ядрышковых организаторов: крупные петли ДНК, активно участвующих в транскрипции рибосомальной РНК;
· Фибриллярный компонент состоит из множества нитей диаметром 5-8 нм, преимущественно во внутренней части ядрышка, и представляет собой длинные молекулы рРНК (первичные транскрипты);
· Гранулярный компонентобразован скоплением плотных мелких гранулярных частиц, представляющие собой зреющие субъединицы рибосом. Рибосомальные субъединицы образуется из рРНК, синтезированной в ядрышке, и белков, синтезированных в цитоплазме.
· Фибриллярный и гранулярный компоненты ядрышка образуют ядрышковую нить – нуклеолонему, которая образует петлистую сеть, выделяющуюся большой плотностью на фоне менее плотного ядерного матрикса
Размеры и объем ядрышек увеличиваются при повышении функциональной активности клетки. Особенно крупные ядрышки характерны для эмбриональных и активно синтезирующих белки клеток, а также клеток быстрорастущих злокачественных опухолей.
Ядрышко исчезает в профазе митоза, в результате инактивации рибосомных генов при конденсации соответствующих хромосом, и вновь формируется в поздней телофазе.
Ядерный матрикс – компонент ядра, в котором располагаются хроматин и ядрышко. Ядерный матрикс образован кариоплазмой и кариоскелетом. Кариоплазма – жидкий компонент ядра, содержащий РНК, ионы, ферменты, метаболиты, растворенные в воде. Кариоскелет состоит из ламины и других фибриллярных белков.
Клеточный цикл – совокупность процессов, происходящих в клетке между двумя последовательными делениями или между её образованием и гибелью. Клеточный цикл включает в себя собственно митотическое деление и интерфазу – промежуток между делениями
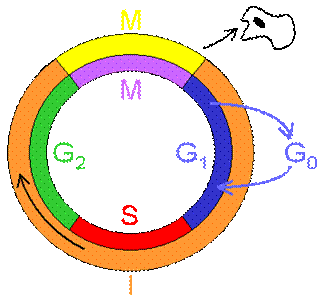
Интерфаза занимает около 90% всего времени клеточного цикла и подразделяется на три периода:
· пресинтетический или постмитотический – G1 (от англ. gap – промежуток);
· синтетический – S;
· постсинтетическийилипремитотический - G2.
Пресинтетический период – G1 – характеризуется активным ростом клетки, синтезом белка и РНК, благодаря чему клетка восстанавливает необходимый набор органелл и достигает нормальных размеров. G1 период длится от нескольких часов до нескольких дней. В течение этого периода синтезируются особые «запускающие» белки – активаторы S периода. Они обеспечивают достижение клеткой точки R (точки ограничения), после которого она вступает в S-период.
Если клетка не достигает точки R, она выходит из цикла и вступает в период репродуктивного покоя (G0).Клетки некоторых тканей под влиянием определенных факторов способны возвращаться из периода G0 в клеточный цикл, клетки других тканей (кардиомиоциты, нейроны) утрачивают эту способность по мере дифференцировки. Абсолютное большинство дифференцированных клеток организма, выполняющих свои специфические функции, не делятся.
Синтетический период –S- характеризуется репликацией (удвоением содержания) ДНК, синтезом гистонов и других белков. В результате происходит удвоение числа хромосом. Одновременно удваивается число центриолей. S-период длится у большинства клеток 8-12 часов.
Постсинтетический период – G2 - длится 2-4 часа и продолжается вплоть до митоза. В течение этого периода запасается энергия, и синтезируются белки, в частности тубулины, необходимые для процесса деления.
Митоз (кариокинез) является универсальным механизмом деления соматических клеток. Во время митоза родительская клетка делится, и каждая из дочерних клеток получает набор хромосом идентичный родительскому, и, таким образом, происходит равномерное распределение генетического материала. Продолжительность митоза – 1-3 часа.
Митоз условно разделяют на 4 основные фазы:
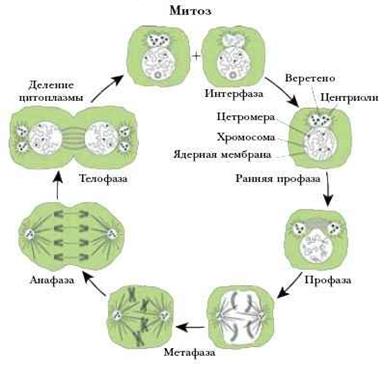
· профазу;
· метафазу;
· анафазу;
· телофазу.
Профазаначинается сконденсации хромосом, которые становятся видимыми в световой микроскоп как нитевидные структуры. Каждая хромосома состоит из двух параллельно лежащих сестринских хроматид, связанных в области центромеры. Ядерная оболочка распадается на мембранные пузырьки и исчезает к концу профазы, так же как и ядрышко. Кариоплазма смешивается с цитоплазмой. Пары центриолей расходятся к противоположным полюсам клетки и дают начало микротрубочкам митотического веретена.
В метафазе хромосомы выстраиваются в области экватора митотического веретена (в равной удаленности от центриолей противоположных полюсов), и образуют картину экваториальной (метафазной) пластинки (вид сбоку) или материнской звезды (вид со сторону полюсов). Сестринские хроматиды к концу этой фазы разделяются щелью, однако удерживаются в области центромеры.
Анафаза начинается с синхронного расщепления всех хромосом на сестринские хроматиды (в области центромеры) и движения дочерних хромосом к противоположным полюсам клеток, происходящего вдоль микротрубочек. Анафаза завершается скоплением на полюсах клетки двух идентичных наборов хромосом, которые образуют картину звезд (стадия дочерних звезд). В конце анафазы начинает образовываться клеточная перетяжка, благодаря сокращению актиновых микрофиламентов, которые концентрируются по окружности клетки.
Телофазахарактеризуется реконструкцией ядер дочерних клеток и завершением их разделения. Ядерная оболочка восстанавливается, хромосомы постепенно деспирализуются, замещаясь картиной хроматина интерфазного ядра, а в конце телофазы вновь появляется ядрышко. Углубление клеточной перетяжки завершается полной цитотомией с формированием двух дочерних клеток. При этом происходит приблизительно равное распределение органелл между дочерними клетками.
Эндомитоз – процесс увеличения числа хромосом внутри ядерной оболочки без последующего деления клетки, что приводит к повышенному содержанию ДНК в ядре – полиплоидии.
Полиплоидные ядра имеют больший объем. Полиплоидные клетки могут также возникнуть вследствие митотического деления без последующей цитотомией. При таком делении образуются двуядерные клетки с увеличенным вдвое набором хромосом. Основной смысл развития полиплоидии заключается в усилении функциональной активности клеток.
Наличие полиплоидных – тетра- (4n, если 1n – гаплоидный набор хромосом) и октаплоидных (8n) клеток – нормальное явление для гепатоцитов (клеток печени), переходного эпителия мочевого пузыря, секреторных клеток поджелудочной и слюнных желез. Уровень полиплоидизации мегакариоцитов красного костного мозга достигает – 16-32n.
По уровню обновления ткани организма подразделяются на три группы – три типа клеточных популяций:
· Обновляющиеся клеточные популяции характеризуются постоянным обновлением. Естественная убыль дифференцированных клеток, специализированных к выполнению определенных функций и неспособных к делению уравновешена образованием новых клеток в результате деления малодифференцированных камбиальных клеток и последующей дифференцировки (физиологическая регенерация). К таким популяциям относят клетки костного мозга и крови, эпителий кишки, эпидермис кожи.
· Растущие клеточные популяции способны к увеличению массы ткани за счет нарастания числа клеток и их полиплоидизации. Их долгоживущие клетки выполняют специализированные функции, но сохраняют способность при стимуляции, под действием некоторых факторов вновь вступать в клеточный цикл, чтобы восстановить свою нормальную численность. К растущим популяциям относят эпителий почек, различных желез, печени.
· Стабильные клеточные популяции состоят из высокоспециализированных клеток с полной потерей способности к делению. К таким популяциям относятся нейроны, кардиомиоциты.
Регуляция клеточного цикла в различных тканях организма осуществляется сложной системой механизмов, стимулирующих или ингибирующих клеточное деление.
Протоонкогены – группа генов-активаторов, контролирующих клеточное деление и дифференцировку. Изменения структуры и усиление активности экспрессии протоонкогенов вызывает развитие опухолей. Повышение активности протоонкогенов может быть связано с изменениями строениями ДНК (в результате мутаций), увеличением количества генов (генной амплификации) или их перегруппировкой, при которой гены размещаются вблизи активного промотора (т.e., участка ДНК, ответственного за инициацию транскрипции). Злокачественная трансформация клетки может возникнуть не только вследствие повышения активности протоонкогенов, но и в результате снижения активности другой группы генов, называемых антионкогенами.
Антионкогены – гены, которые продуцируют супрессоры опухолевого роста, угнетающие митотическую активность клеток. Пример антионкогенов – ген р53. Ген р53 обеспечивает поддержание стабильности генетического аппарата и контролирует клеточный цикл. Его экспрессия резко усиливается при повреждении ДНК. Активация гена р53 приводит к остановке клеточного цикла (выходу в G0 ) для репарации ДНК, а при тяжелых повреждениях запускает программу апоптоза (клеточной гибели). Выявлена четкая связь между утратой функции гена р53 (в результате мутации или делеции) и развитием более 50 видов злокачественных опухолей у человека.
Факторы ростапредставляют собой гликопептиды, продуцируемые клетками различных тканей, усиливающие митотическую активность в определенных клетках-мишенях, имеющих специфические рецепторы на плазмолемме. К ним относятся фактор роста нервов, инсулиноподобные факторы роста, колониестимулирующие факторы, интерлейкины и другие цитокины.
Кейлоны, напротив, подавляют клеточное деление. Кейлоны образуются всеми зрелыми дифференцированными клетками и локально воздействуют на камбиальные элементы (стволовые и полустволовые клетки) этой же ткани. Они обеспечивают стабильную численность клеточной популяции, а их выделение контролируется механизмом отрицательной обратной связи. При уменьшении численности зрелых клеток данной популяции (например, потеря лейкоцитов при кровотечении или эпидермиса при ранении) продукция кейлонов снижается, что приводит к усилению митотической активности клеток, способных к делению, - репаративной регенерации.
Число клеток в организме, органах и тканях регулируется гомеостатическими механизмами и динамическим равновесиеммеждуобразованием клетоки их гибелью. Гибель клеток, наряду с их размножением (пролиферацией) и дифференцировкой, является одним из ключевых процессов в обеспечении нормальной жизнедеятельности различных тканей
При гибели клеток могут наблюдаться два вида морфологических изменений, которые соответствуют различным механизмам её развития:
· некрози
· апоптоз.
Некрозвозникает под действием резко выраженных повреждающих факторов: перегревания (гипертермии), переохлаждения (гипотермии), недостатка кислорода (гипоксии), нарушения кровоснабжения (ишемии), метаболических ядов, механической травмы и др. Некроз – «смерть в результате несчастного случая».
Поздние явления при некрозе включают: разрыв ядерной оболочки, плазмолеммы и мембран органелл, разрушение и растворение ядра (кариолизис), исчезновение клеточных границ и распад клетки
Апоптоз - физиологическая (запрограммированная)гибель клеток; «смерть клетки в результате самоубийства (самоуничтожения)». Апоптоз – это активный, генетически контролируемый процесс, регулируемый внутренней программой, которая запускается внешними факторами. Факторы могут разными: повреждающие физические и химические факторы, умеренные по интенсивности; некоторые инфекции (вирусные); воздействие физиологических активаторов (индукторов) апоптоза; дефицит стимулирующих факторов, потеря контакта с другими клетками и др.
Развитие апоптоза индуцируется особыми генами (киллерными генами – р53 и др.). Это энергоёмкий процесс и сопровождается активацией сигнальных систем в клетке. Развитие апоптоза морфологически на светооптическом уровне проявляется уплотнением ядра (кариопикноз и кариорексис без разрушения кариолеммы), конденсацией цитоплазмы, которая уплотняется, сморщивается и уменьшается в размерах, органеллы при этом сохраняют свою целостность. При прогрессировании апоптоза изменяется форма клетки – образуются многочисленные крупные вздутия и выросты на поверхности – и происходит распад клетки на фрагменты – апоптозные тела.
IY. МЕЖКЛЕТОЧНЫЕ СОЕДИНЕНИЯ.
МЕЖКЛЕТОЧНЫЕ СОЕДИНЕНИЯ
Внешние клеточные мембраны участвуют в образовании межклеточных контактов, которые обеспечивают межклеточные взаимодействия.
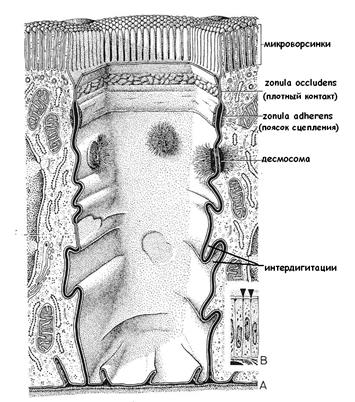
Простое межклеточное соединение – сближение плазмолемм соседних клеток на расстояние 15-20 нм. Важную роль при этом играют клеточные рецепторы-гликопротеины, называемые клеточными адгезионными молекулами (КАМ), такие как кадгерины, интегрины, способные распознавать и связывать плазмолеммы соседних клеток. Интегрины – трансмембранные белки, внутриклеточная молекула интегрина через ряд других промежуточных белков (таких как винкулин, α-актинин) связана с цитоскелетом. Наружная часть молекулы через другие гликопротеины (фибронектин, ламинин) связана с клетками и молекулами внеклеточного матрикса. При этом плазмолеммы соседних клеток могут формировать интердигитации, то есть взаимные выпячивания двух соседних клеток. Такой тип межклеточных соединений усиливает механическую прочность соединения клеток и увеличивает площадь обменной поверхности.
Сложные межклеточные соединения – небольшие парные специализированные участки плазматических мембран соседних клеток. Сложные межклеточные соединения подразделяются на изолирующие (запирающие), сцепляющие, обусловливающие механическое сцепление и соединение клеток, и коммуникационные соединения, обеспечивающие химическую (метаболическую, ионную) и электрическую связь между клетками. Особенно ярко выражены межклеточные соединения в эпителиальных тканях (рис. 13).
К изолирующим соединениям относятся плотные контакты.
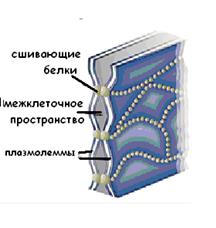
Плотный контакт (zonula occludens) окружает апикальную часть клеток по периметру в виде пояска. Это область частичного слияния наружных листков плазмолемм двух соседних клеток. Специальные белки, образующие подобие ячеистой сети (окклюдины), как бы «сшивают» соседние плазмолеммы. Основная функция плотного контакта – блокировать проникновение и распространение веществ по межклеточному пространству.
К сцепляющим (заякоривающим) соединениям относят поясок сцепления и десмосомы. Для сцепляющих соединений характерно наличие слоя примембранных белков, примыкающих к цитоплазме в области контакта, к которым подходят фибриллярные элементы цитоскелета. Поясок сцепления (zonula adherens, опоясывающая десмосома) также опоясывает клетки в виде ленты, но локализуется на латеральной поверхности клеточной мембраны ниже, чем плотный контакт. Здесь клетки связаны друг с другом интегральными гликопротеидами, к которым примыкает слой примембранных белков (винкулин и др.). С этим слоем связаны пучки актиновых микрофиламентов. Десмосома (macula adherens)- парная структура, состоящая из утолщенных и уплотненных участков цитоплазмы, прилегающих к плазмолеммам соседних клеток, так называемых пластинок прикрепления, разделенных межклеточной щелью. Каждая пластинка прикрепления имеет форму диска (диаметр около 0.5 мкм) и содержит особые белки (десмоплакины и др.), к которым прикреплены пучки промежуточных филаментов (тонофиламентов). При этом находящиеся в межклеточном пространстве Са2+-связывающие белки взаимодействуют с пластинками прикрепления, благодаря чему усиливается механическое сцепление клеток. Десмосомы не имеют определенной локализации и разбросаны по поверхности клетки.
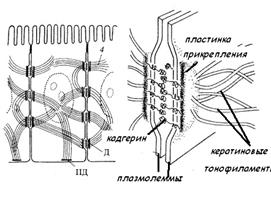
Десмосома
Рис.13.
Коммуникационные соединения представлены щелевыми контактами и синапсами. Щелевое соединение (нексус) представляет собой участок протяженностью 0,5-3 мкм, где плазмолеммы разделены узкой межклеточной щелью (2-3 нм). При этом в структуре плазмолемм соседних клеток друг против друга располагаются трубчатые трансмембранные структуры – коннексоны (из белка коннексина), которые образуют межцитоплазматические каналы, обеспечивающие свободный обмен низкомолекулярными соединениями между клетками (рис.14). Число конексонов в одном щелевом контакте обычно исчисляется сотнями. Функциональная роль щелевых соединений заключается в переносе ионов и мелких молекул от клетки к клетке.
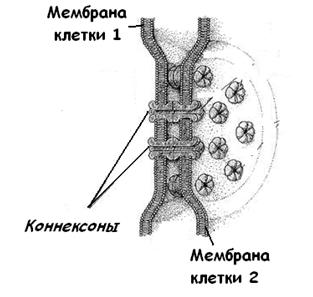
Рис.14. Щелевой контакт
(нексус)
Синаптические соединения – высокоспециализированные контакты нервных клеток, проводящие импульсы в одном направлении. Синаптические контакты устанавливаются также между нейронами и мышечными и железистыми клетками.
Дата добавления: 2016-03-22; просмотров: 2832;