Можно ли считать ДВП памятью?
Долговременная потенциация в гиппокампе, несомненно, представляет собой необычайно перспективную модельную систему, которая уже позволила получить много данных о том, как можно связать нейрофизиологические изменения с биохимическими и структурными механизмами. Однако я не убежден, что уже поставлены наиболее важные биохимические вопросы, - отчасти потому, что слишком много внимания уделялось интимным синаптическим процессам, связанным с инициацией ДВП, и на удивление мало - самой долговременности этого феномена, которая кажется мне наиболее интересной его стороной. Изменения в притоке ионов кальция, в фосфорилировании мембранных белков или в активации NMDA-рецепторов - все это возможные механизмы кратковременного изменения электрических свойств клетки, но если мы хотим, чтобы ДВП действительно служила моделью долговременной памяти, важно знать, что сообщает этому изменению устойчивость, что привносит букву Д в аббревиатуру ДВП.
Особая ценность ДВП как модели, помимо возможности еще более свободно, чем в случае с аплизией, переходить с одного уровня анализа на другой (от интактного организма к срезу), определяется геометрическим фактором. Клеточные и биохимические изменения, которые нужно связать с поведенческими реакциями, так же как образование следов памяти, должны быть точно локализованы во времени и пространстве, в соответствии с критериями, сформулированными в начале этой главы. Гиппокамп - это одна из областей мозга млекопитающих, строение, связи и геометрия которой хорошо известны, что делает в принципе осуществимым такое картирование. Поэтому при разработке моделей памяти гиппокамп открывает большие возможности для проверки хеббовских закономерностей обучения на нейронах, связи которых действительно известны, а не остаются предметом догадок [26}.
Возвращаясь к вопросу, с которого я начал обсуждение феномена долговременной потенциации, попробуем решить, действительно ли она может служить моделью длительных изменений в нервной системе или, более того, это и есть сам механизм памяти? В пользу того, что ДВП - это механизм, обеспечивающий хранение в мозгу следов памяти, говорит (как я уже отмечал) прежде всего хорошо известная роль гиппокампа в различных формах памяти, так же как и факт существования ассоциативных форм ДВП. Но дальнейшие аргументы носят уже гипотетический характер. Так, например, ДВП усиливается у крыс, обученных находить корм в оперантных задачах, тогда как блокирующие ДВП вещества препятствуют также обучению в условиях водного лабиринта. С возрастом крысы утрачивают способность обучаться новым навыкам, и в то же время у них не наблюдается потенциация. Но если сопоставить эти доводы с критериями, сформулированными в начале главы, они уже не будут выглядеть столь убедительными.
Имеются два рода наблюдений, которые, мне кажется, ставят под сомнение правомерность прямых параллелей между ДВП и памятью. Во-первых, появляется все больше данных, что подобные ДВП явления не ограничены гиппокампом - при известных обстоятельствах они проявляются во многих других областях мозга, в том числе (и особенно) в его коре, как показали Линн Байндмен в Лондоне и Алексей Воронин в Москве [27]. Это ставит под сомнение доводы в пользу связи между ДВП и процессами памяти в гиппокампе. В одном из своих изящных исследований Кэрол Барнс, чьи ранние работы были посвящены выявлению корреляций между старением, научением и ДВП, содержала крыс в информационно-обогащенной среде, как это было в экспериментах Розенцвейга, Беннета и Даймонда (глава 6). Она нашла, что чем дольше крысы оставались в такой среде, тем слабее у них в последующем индуцировалась ДВП в гиппокампе [28].
Моя собственная интерпретация этого наблюдения (не обязательно совпадающая с интерпретацией автора) подсказывает мысль, что ДВП могла быть не чем иным, как простым артефактом, возможным только тогда, когда животные выросли в сильно обедненных условиях лаборатории. Может быть, способность их к формированию когнитивных карт в гиппокампе в таких условиях просто не находила себе применения, и поэтому их мозг жадно реагировал на новую информацию при нейрофизиологической стимуляции входных нервных путей. В более нормальных, естественных условиях, где такая стимуляция и ее последствия были бы повседневными событиями, ДВП, возможно, вообще бы отсутствовала. Таким образом, ДВП может быть артефактом, следствием обедненных условий содержания лабораторных животных. Это сам по себе интересный феномен, но еще более аномальный, чем позволяло предполагать осторожное предупреждение Блисса и Лёмо в 1973 году. Прежде чем дальше теоретизировать по поводу такого предположения, его нужно проверить, чтобы установить, можно ли вызвать ДВП у животных, выросших в естественных, не столь однообразных условиях дикой природы.
Глава 10
Никто кроме нас, цыплят
Наконец-то. Шесть глав на отступления, и я снова могу вернуться к цыплятам. Прошу извинить меня - это было долгое путешествие, но я не сумел найти путь короче. Еще несколько абзацев, и мы у цели. 1977-й год был первым за всю мою жизнь, когда я был свободен от лекций, и я провел пару месяцев в Австралийском национальном университете в Канберре. Я выбрал Канберру, потому что там работала с курами большая группа исследователей и в их числе один из моих бывших учеников. К тому времени я несколько разочаровался в изучении импринтинга, которое в моей лаборатории уже несколько лет было привычной рутиной, и мне хотелось поискать иную форму научения у цыплят. Группа в Канберре начала изучать действие различных препаратов на образование следов памяти у кур с помощью нового метода, модифицировав тест, предложенный старейшим нейробиологом из Лос-Анджелеса Артом Черкином.
Данное им описание разработки этого метода в середине шестидесятых годов очень характерно для Черкина. Он пишет, как, наблюдая за молодыми курами, заметил, что они обследовали окружающий мир, склевывая крошки и другие мелкие предметы, включая даже собственные экскременты, но быстро научались отличать съедобное от несъедобного. «Черт бы меня побрал, - продолжает он, - если мне приходило в голову, что мою систему будут называть дерьмовыми экспериментами Черкина». Итак, Черкин вместе с Илей Ли-Тенг натолкнулся на мысль предлагать цыплятам клевать маленькие разноцветные бусины. Если бусина имела горький вкус, так как исследователи смочили ее спиртом, хинином или едким метилантранилатом, цыплята, однажды клюнув ее, выражали отвращение, энергично тряся головой и вытирая клюв о пол клетки, а когда через несколько секунд или суток им снова предлагали сухую бусину такого же цвета, они всякий раз пятились назад и отказывались клевать. Такова была в своей основе модель разового обучения пассивному избеганию, которой я заинтересовался.
Обучение было разовым, так как цыпленку достаточно было клюнуть бусину однажды, чтобы извлечь урок. Это была выработка реакции избегания, поскольку птенец переставал делать то, что делал бы в иных условиях. Это было пассивное избегание, так как цыпленок должен был лишь воздерживаться от контакта, не предпринимая никаких активных действий, чтобы избежать неприятного ощущения.
Импринтинг был весьма впечатляющей формой научения, но, с моей точки зрения, имел тот недостаток, что для его формирования птенца приходилось часа два подвергать воздействию раздражителя (проблесков света и т. п.). В этот период запоминание происходило медленно, и связанные с ним клеточные изменения смешивались с эффектами зрительной стимуляции. Пэт, Габриел и я потратили несколько лет, пытаясь разделить эти факторы, но модель импринтинга лучше было оставить, если мы действительно хотели изучать те клеточные процессы в первые минуты после прекращения стимуляции, которые отражали разные стадии формирования памяти1.
*1) Еще одно замечание относительно импринтинга. Когда бы мы ни докладывали наши результаты в присутствии психологов, кто-нибудь обязательно спрашивал, применимы ли представленные данные к другим формам научения, поскольку «импринтинг - это уникальное явление». Пэт Бейтсон, как этолог, всегда давал исчерпывающие разъяснения [1], и мы не видели оснований считать, что клеточные механизмы здесь существенно отличаются от механизмов любой другой формы научения у позвоночных. Однако выработка пассивного избегания - это такая задача, которая больше устраивает психологов, хотя впоследствии я понял, что возможность обучения за один сеанс, которую я считал преимуществом метода, некоторых из них смущала, так как не позволяла количественно оценивать выполнение задачи с такой же точностью, как при обучении в десятках и сотнях проб. Правда, есть люди, которым трудно угодить.
Этой цели отвечала выработка пассивного избегания, хотя бы потому, что она определялась в одноразовом тесте. Кроме того, она была предпочтительна и по чисто практическим соображениям. Для импринтинга и последующей оценки его эффективности нужен громоздкий и сложный аппарат, в котором одновременно можно было обучать лишь нескольких птенцов. Это не имело особого значения при физиологических или анатомических исследованиях, когда все равно приходится работать лишь с небольшим числом животных, но сильно тормозило биохимические исследования, где их требуется гораздо больше. Для опытов с пассивным избеганием нужен был лишь набор небольших (20x30 см) клеток всего на двух цыплят каждая, которые я упоминал в главе 2 (в США для этой цели используют даже коробки из-под молока объемом в кварту). А само тестирование состояло в простом повторном предъявлении цыплятам бусины в течение определенного времени, скажем десяти секунд, и регистрации их реакций.
Когда я приехал в Канберру, там уже не было Мэри Гиббс, организовавшей там работу с пассивным избеганием, - она перебралась в Ла-Троуб, городок Мельбурнского университета в нескольких сотнях километров от прежнего места работы. Ее больше всего интересовала динамика формирования памяти во времени, и для выделения отдельных фаз этого процесса (кратковременной, долговременной и промежуточной памяти) она использовала множество различных веществ, в том числе агенты, блокирующие поступление в клетки калия и других ионов, а также ингибиторы белкового синтеза. По ее наблюдениям, для каждой фазы была характерна чувствительность к веществам определенного типа. Самая ранняя фаза длилась всего несколько минут после сеанса обучения, промежуточные фазы занимали не более часа, а ингибиторы белкового синтеза не оказывали действия, если вводились больше чем через час после тренировки (рис. 10.1)1.
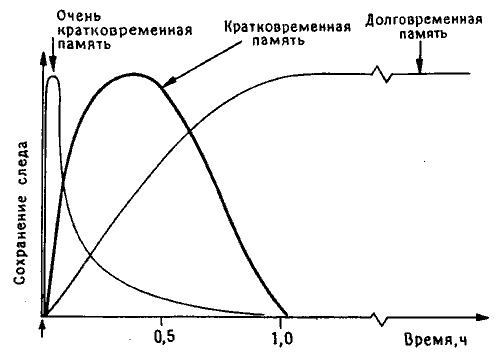
Рис. 10.1. Динамика следов памяти у цыпленка. Мэри Гиббс предложила выделять в этом процессе три фазы (впоследствии она добавила четвертую, промежуточную фазу, которая здесь не показана).
*1) В этом методе используется радиоактивное соединение, которое количественно связывается с рецепторами ацетилхолина, так что связанная радиоактивность пропорциональна количеству рецепторов. Это соединение, известное под шифром QNB, было нетрудно достать в других лабораториях, но мои коллеги никак не решались сказать мне, откуда они получали этот препарат. Только потом я узнал причину: препарат был разработан как потенциальное химическое оружие и поступал через Портон-Даун (первоначально он обозначался BZ и испытывался Корпусом химической защиты США во время вьетнамской войны как средство дезориентации, которое при распылении над вражескими позициями лишало противника желания и сил бороться). Неудивительно, что коллеги проявляли такую скрытность, учитывая мою широко известную оппозицию исследованиям в области химического и биологического оружия, хотя я всегда очень скептически относился к возможности практического применения такого вещества в военных целях. Этот вопрос снова возник в 1991 году как следствие войны в Персидском заливе, когда предметом общего беспокойства стали запасы химического оружия у Ирака. Оказалось, что в течение нескольких лет британские компании поставляли в эту страну огромные количества QNB под предлогом его необходимости для борьбы с кишечными инфекциями в иракской армии!
Меня, однако, интересовала биохимия, а не фармакология формирования памяти. Мы договорились с Мэри, что она будет обучать цыплят в Ла-Троубе в точном соответствии с протоколом, который она сама разработала на основе метода Черкина и который мы фактически используем до сих пор, как я описывал в главе 2. После этого она должна была замораживать пробы мозга под соответствующими шифрами и пересылать их мне самолетом, так как более удобной связи с Канберрой не было. Мне же предстояло разделять мозг на те же области, которые мы привыкли грубо различать при изучении имприн-тинга, и проводить биохимические анализы. В то время я был особенно увлечен выяснением роли в формировании памяти одного из важнейших нейромедиаторов - ацетилхолина - и наладил простой и быстрый метод определения его мускариновых рецепторов, отличающихся от других типов рецепторов ацетилхолина. Я уже знал, что крыша переднего мозга кур особенно богата такими рецепторами, а мой метод позволял достаточно быстро измерять их содержание в двух сотнях проб за 12-часовой рабочий день (хотя потом, конечно, требовалось еще несколько дней для анализа результатов).
Согласно нашему плану, Мэри должна была обучать цыплят не клевать горькую бусину, смоченную метилантранилатом, и одновременно тренировать другую группу цыплят клевать бусину, смоченную водой. Последние при дальнейшем тестировании клевали сухую бусину и, таким образом, служили контролем, поскольку у них не была выработана реакция избегания. (Если вы следили за логикой изложения в предшествующих главах, вы сразу заметите, что это не лучший контроль, так как цыплята, обученные клевать бусину, смоченную водой, могли что-то знать и о других ее свойствах. Но я вернусь к этому вопросу позднее. В то время нам казалось, что будет несложно провести эксперимент, спланированный описанным образом.) Мне предстояло определять количество мускариновых рецепторов в различных отделах мозга цыплят в разные сроки после того, как они клевали бусины, смоченные метилантранилатом или водой, чтобы выявить кратковременные или длительные изменения их содержания.
Чтобы выполнить запланированную работу в полном объеме, времени, казалось, было вполне достаточно. Были, однако, задержки с налаживанием метода в незнакомой лаборатории; потом я галопом объехал около десятка университетских городков, где провел семинары, и в результате лишь в последние несколько дней пребывания в Австралии, во время долгого автомобильного переезда по необжитой местности на конгресс в Брисбейне, мне удалось расшифровать и суммировать все полученные данные. Я с радостью обнаружил, что через 30 минут после того, как цыплята клевали горькую бусину, содержание мускариновых рецепторов возрастало в той же самой области мозга, где мы наблюдали изменения при импринтинге. Но это повышение носило временный характер и сохранялось не более трех часов после обучения. Это, по-видимому, означало, что увеличение количества мускариновых рецепторов было связано с ранними фазами формирования следов памяти.
Конечно, этот вывод нельзя было считать окончательным, что сразу становилось ясно при обращении к моим критериям, изложенным в предыдущей главе. Например, наблюдавшееся увеличение могло быть реакцией на вкус самого метилантранилата, а не следствием сформировавшейся ассоциации между клеванием бусины и ее вкусом. Уже после моего возвращения домой Мэри проверила такую возможность довольно примитивным способом: она залепляла цыплятам глаза и вкладывала в клюв комочек ваты, смоченный метилантранилатом. В этом случае не было повышенного связывания нейромедиатора с мускариновыми рецепторами. Однако я все еще соблюдал осторожность и дождался, когда спустя несколько месяцев Мэри нанесла ответный визит в Англию. К тому времени я воспроизвел использованную ею процедуру тренировки в моей лаборатории, и мы смогли с самого начала повторить весь эксперимент. Результат был тот же самый (хотя на этот раз эффект оказался несколько слабее), и мы опубликовали первую статью, основанную на данных, полученных в 1980 году [3].
Благодаря успешным опытам с мускариновыми рецепторами я стал приверженцем модели пассивного избегания до такой степени, что почти все другое практически исключалось. Мы разобрали оборудование для исследования импринтинга, чтобы занять освободившееся место клетками для изучения пассивного избегания, и стал искать источники финансирования, чтобы полным ходом развернуть новую работу. Начиная с 1980 года лаборатория трудится исключительно над разработкой этой проблемы.
Я мог бы многое порассказать о прошедшем десятилетии, отданном этой работе. Кое-что встает со страниц научных публикаций - этих необычных, словно скованных произведений письменности, столь же строгих по форме, как сонет, а по содержанию - четко спланированных, безукоризненно проведенных, убедительных наблюдений, опирающихся на более ранние результаты и позволяющих в конце указать направление будущих экспериментов («необходимы дальнейшие исследования, чтобы...»). Другой рассказ - о том, что стоит за этими статьями, хроника экспериментов в их реальной последовательности. Она отразила бы наши метания от одной задачи к другой, от одного метода к другому, от биохимии к поведению (ибо наша лаборатория в большей степени, чем все другие мне известные, словно сорока подбирает разные методы: нам подходит любой из них, если только а) он нам по средствам, б) он помогает нам двигаться вперед в главном направлении, и в) мы знаем, как им пользоваться, или можем найти знающих людей). Некоторые сами собой напрашивающиеся важные вопросы приходилось на многие годы оставлять без ответа, так как я не мог найти к ним подхода, не имел времени для проведения экспериментов или не знал, где найти средства для приобретения нужных реактивов и оборудования. Другие возникали и решались по обстоятельствам, когда в лабораторию приезжал специалист или появлялся диссертант, владевший нужным методом или имевший идею, способную помочь нашему продвижению в том направлении, которое без этого могло бы и не привлечь нашего внимания. Наконец, некоторые мысли приходили в голову где-нибудь по дороге домой после случайно прочитанной статьи или услышанного на конференции разговора. Иногда, когда становились известны результаты, полученные в другой лаборатории, приходилось менять план и задачу уже начатого эксперимента. Все это, как много лет назад заметил Питер Медавар в своем классическом эссе «Не надувательство ли научная статья?» [4], - неотъемлемые атрибуты исследовательской работы, которые в окончательном отчете о ней, будь то журнальная публикация или обзор, предстают в рафинированном виде, в значительной степени, хотя и не полностью, отфильтрованными, как при обсуждении данных по аплизиям и ДВП в предыдущей главе. Я никому, кроме себя, не пожелал бы работать таким образом!1 1
*1) Но есть и еще одна, более личностная сторона дела, чем хронологическое перечисление событий. Это рассказ о людях и взаимодействии между ними в научном коллективе, о том, кто мы такие, во что одеваемся, как выглядим, кто и с кем находится или находился в тех или иных отношениях. Психодинамика небольшой исследовательской группы, связанной общими научными интересами и работающей в университетском городке, довольно далеко от крупных центров, имеет свою привлекательность: тут и соперничество между членами группы и с другими такими же группами, и бесконечные стычки из-за субсидий и помещений, и чередование преподавательской работы с научной... Это обширный материал для целого романа, пожелай им кто-нибудь воспользоваться; ведь речь идет, скажем, о двадцати пяти довольно молодых людях из разных стран, временно (от месяца до трех лет) работающих в данном коллективе. (Ч. П. Сноу много лет назад попытался написать об этом, но мог бы и не нести такую несусветную чушь, потому что хорошо знал то, о чем рассказывал.) Сам же я еще не готов к этому.
В этой и следующей главах я собираюсь поведать две разные истории. Первая не хронологическая, а логическая: о том, как, выработав критерии, с которых я начал главу 8, я хотел найти удовлетворяющие им явления в опытах с выработкой пассивного избегания. Я надеюсь, что этот рассказ будет выглядеть убедительно, так как именно его я пытался преподнести моим коллегам-нейробиологам, но в одном важном смысле он будет не до конца объективен. Не потому, что я намеренно искажаю или неверно излагаю наши результаты, а потому, что отобрал и систематизировал их в соответствии с моими теоретическими установками и творческими задачами - это окрасит действительность в те цвета, которым я отдаю предпочтение. Вторая история совсем иная - в ней не будет четких выводов, ибо она еще не закончена и грозит снова превратить в хаос порядок, установленный мною в первом рассказе. Но она, может быть, позволит лучше представить всю неопределенность и непредсказуемость экспериментальных исследований, чем предшествующее ей гладкое изложение событий.
Но довольно преамбул и манифестов. Пора приступить к первому рассказу.
Дата добавления: 2016-03-05; просмотров: 700;