Нормальные и аномальные производные гемоглобина
Процесс взаимодействия молекулы кислорода с гемоглобином можно описать с помощью следующего уравнения (рис. 4):

Рис. 4. Присоединение кислорода к гемоглобину
В результате присоединения кислорода к катиону Fe2+ гема образуется утворюється оксигемоглобин (рис. 5).
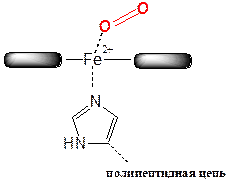
Рис. 5. Схема присоединения молекулы кислорода к гемму с образованием оксигемоглобина.
Процесс образования оксигемоглобина сопровождается изменением конформации его полипептидных цепей. Наиболее выраженные сдвиги происходят в области положения гема. В начале, в структуре дезоксигемоглобина атом железа выдвигается за плоскость протопорфиринового кольца в сторону положения остатка гистидина (рис. 6). Затем, при присоединении кислорода происходит втягивание атома железа в плоскость кольца. Это способствует изменению пространственной укладки полипептидной цепи, связанной с гемом. В результате подобных конформационных сдвигов в молекуле оксигемоглобина изменяется характер взаимодействия полипептидных цепей между собой. Молекула оксигемоглобина становится более компактной, чем молекула дезоксигемоглобина.
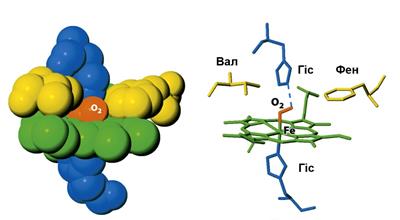
Рис. 6. Изменение конформации полипептидной цепи гемоглобина в результате присоединения кислорода к гему (Berg J.M. et al., 2006)
Изменение конформации полипептидных цепей при присоединении кислорода сопровождается изменением спектральных свойств белковой молекулы, а также её цвета: раствор дезоксигемоглобина имеет темно-красный цвет, а раствор оксигемоглобина - ярко-красный. Это явление объясняет отличие цвета венозной крови (обогащена дезоксигемоглобином) от цвета артериальной крови (обогащена оксигемоглобином).
Конформационная перестройка полипептидной цепи, возникающая при присоединении к гему одной молекулы кислорода, способствует резкому повышению сродства других трёх гемов молекулы гемоглобина к кислороду, облегчая его присоединение.
Процесс образования оксигемоглобина является обратимым и находится под контролем многих факторов. Особое значение среди них имеют концентрации H+, CO2, Cl- и 2,3-дифосфоглицерата. При их возрастании снижается способность гемоглобина связывать кислород.
Оксигемоглобин является нестойким соединением, который распадается при снижении концентрации кислорода в окружающей среде. По этой причине образование оксигемоглобина происходит преимущественно в капиллярах легких, для которых характерно высокое парциальное давление кислорода. При перемещении эритроцита из легких к другим (периферическим) тканям внутренних органов оксигемоглобин разрушается с высвобождением кислорода, так как парциальное давление кислорода здесь гораздо меньше, чем в легких.
Процесс распада оксигемоглобина с высвобождением кислорода в периферических тканях усиливается при повышении в них концентрации H+ (здесь рН ниже, чем в легких) и содержания в них углекислого газа. Последнее явление обусловлено в них интесивно протекающими окислительно-восстановительными процессами, связанными с тканевым дыханием.
Регуляторное влияние углекислого газа и Н+ на связывание и высвобождение кислорода гемоглобином объясняется эффектом Бора. В основе механизма эффекта Бора лежит явление обратимой взаимосвязи между процессами связывания кислорода и высвобождения Н+ гемоглобином. Свойство гемоглобина обратимо связывать кислород и направленно транспортировать его от легких к тканям внутренних органов зависит от величины парциального давления кислорода (рО2) в окружающей среде (рис. 7).
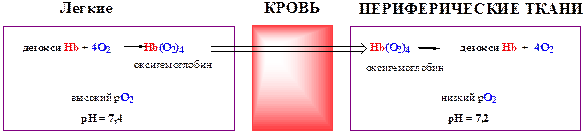
Рис. 7. Направленный транспорт кислорода между легкими и периферическими тканями.
Следующим производным гемоглобина является карбгемоглобин, который образуется при взаимодействии Нb с углекислым газом. В данном соединении СО2 присоединяется не к катиону железа Fe2+, а к NH2-группе цепей глобина:

Таким образом из тканей организма к легким транспортируется до 10 - 15 % СО2. Реакция обратима - образование карбгемоглобина происходит в тканях организма, а его распад - в легких.
Ещё одним производным гемоглобина является карбоксигемоглобин (HbCO) - соединение гемоглобина с угарным газом (монооксидом углерода). Данное соединение более стойкое, чем оксигемоглобин. Поэтому при одновременном вдыхании смеси кислорода и угарного газа преимущественно образуется карбоксигемоглобин:

То есть высокотоксичный угарный газ имеет большее сродство к гемоглобину, чем кислород. Реакция образования HbCO может стать обратимой только при условии наличия высоких концентраций кислорода. Если в воздухе содержится от 0,05 % до 1 % СО, то 95 % гемоглобина переходит в форму HbCO. Тяжёлое отравление угарным газом наступает тогда, когда в воздухе более 0,1 % СО.
При действии окислителей в гемоглобине может происходить окисление катиона Fe2+ в катион Fe3+, что сопровождается образованием метгемоглобина (единственный Hb, содержащий катион Fe3+). Метгемоглобин также не способен связывать и переносить кислород, то есть его физиологическое действие аналогично действию карбоксигемоглобина. Незначительная концентрация метгемоглобина более безопасна, чем такая же концентрация HbCO. Поэтому некоторые стимуляторы образования метгемоглобина используют как антидоты для лечения отравлений цианидами. Метгемоглобин может связывать до 30 % смертельної дозы HCN с образованием малотоксичного соединения цианметгемоглобин. Образованию метгемоглобина способствуют вещества: метиленова синь, натрия нитрит и другие окислители, способные превратить катион Fe2+ гема в катион Fe3+, что сопровождается преходом красного окрашивания раствора в коричневый (при кислых значениях рН).
В настоящее время для выявления отравления СО или образования метгемоглобина в крови пациентов используют спектральный анализ крови в диапазоне длин волн излучения 400-900 нм. Полосы поглощения HbO2 и HbCO имеют сходство и расположены соответсвенно в желтой и зеленой частях спектра, но при этом, у HbCO они смещены в сторону более коротких длин волн. Полоса поглощения метгемоглобина расположена в красной части спектра.
Миоглобин
Миоглобин является одним из важных белков представленных в составе клеток скелетной мускулатуры и миокарда человека. Его основная функция связана с депонированием кислорода в мышцах. Очень богаты миоглобином скелетные мышцы морских животных, которые проводят много времени в движении под водой. Большое содержание миоглобина позволяет этим животным запасать значительную концентрацию кислорода, и тем самым, обеспечивать жизнедеятельность при длительном нахождении без кислорода под водой.
Молекула миоглобина состоит из одной полипептидной цепи, содержащей 153 аминокислотных остатка, и связанного с ней одного гема. Полипептидная цепь в пространстве образует глобулярную структуру, внутри которой в гидрофобном «кармане» помещен гем (рис. 8). Присоединение гема к полипептидной цепи осуществляется через атом азота гистидинового остатка полипептидной цепи. Стабилизация связи простетической группы (гема) и полипептидной цепи происходит также за счет взаимодействия тетрапиррольного кольца гема с неполярными аминокислотными радикалами, которые формируют гидрофобную полость.
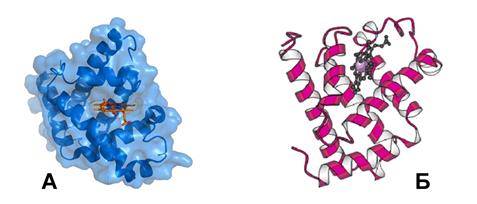
Рис. 8. Трехмерная структура молекулы миоглобина – А (красным цветом обозначено положение гема в молекуле) и её модель – Б
(Berg J.M. et al., 2006)
Среднее содержание миоглобина (Mb) составляет 0,3% от массы тела взрослого человека и повышается в мышечной ткани при дительных физических нагрузках. Приблизительно также, как описывалось образование молекулы оксигемоглобина, происходит обратимое связывание молекулы кислорода с катином железа гема за счет шестой координационной связи с образованием молекулы оксимиоглобина (рис. 9).
Миоглобин связывает кислород О2 в 5 раз быстрее, чем гемоглобин и создает запас кислорода в мышце. Подобно гемоглобину миоглобин образует производные с угарным газом и с цианидами. За счет миоглобина мышцы приобретают красный цвет, а сам Mb (по данным спектральных исследований) имеет широкую полосу поглощения при длине волны 564 нм. Количество кислорода, которое связывается с миоглобином («процент насыщения»), зависит от концентрации кислорода в среде, окружающей миоглобин (эту концентрацию выражают как рО2 – парциальное давление кислорода).
В условиях кислородного голодания (например, при высокой физической нагрузке) кислород освобождается из комплекса с миоглобином и поступает преимущественно в митохондрии мышечных клеток. Где осуществляется синтез АТФ за счет окислительного фосфорилирования.
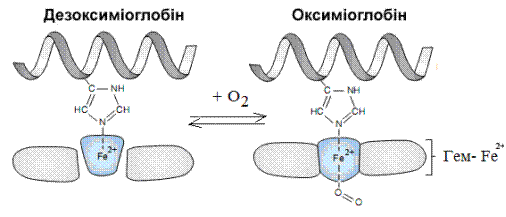
Рис. 9. Механизм присоединения кислорода к молекуле миоглобина (Smith C. et al., 2005, с изм.)
Цитохромы
Среди гемопротеинов особое место занимают цитохромы. Они входят в состав цепей транспорта электронов митохондрий, эндоплазматического ретикулума и хлоропластов (у растений). Наличие простетической группы гема с катионом железа, который способен менять степень окисления, обеспечивает их участие в транспорте электронов по цепи. Большинство цитохромов подобно миоглобину содержат одну молекулу гема и одну полипептидную цепь.
Известно около 30 разных цитохромов, которые по спектрам поглощения делятся на группы a, b, c, d (рис. 10). Гемы этих цитохромов также являются производными протопорфирина IX. Особенности структуры гема обусловливают отличия в проявлении оптических свойств цитохромов и в значениях их редокс-потенциалов.
Кроме структуры боковых радикалов порфиринов, цитохромы отличает друг от друга строение белковой части и способ присоединения гема белков. В различных типах цитохромов гем по-разному соединяется с полипептидной цепью. Например, цитохромы типа с, в отличие от других, содержат прочно связаный с аполипопротеином гем. Он ковалентно присоединяется за счет двух виниловых радикалов к сульфгидрильным группам цистеиновых остатков полипептидной цепи (рис. 10).

Рис. 10. Типы гемов, входящих в структуру цитохромов.
Цитохром с является компонентом дыхательной цепи митохондрий. Ниже представлена модель цитохрома с, созданная на основе детального изучения его третичной структуры (рис. 11).
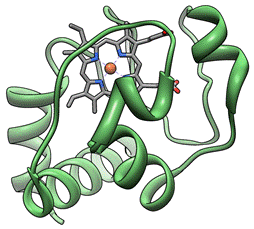
Рис. 11. Модель цитохрома с (Bushnell et al., 1990)
В эндоплазматическом ретикулуме печени содержится еще один широко распространенный цитохром - цитохром Р450, названный так потому, что впервые был открыт в Филадельфии (Philadelphia), США, а комплекс его восстановленной формы с угарным газом СО имеет максимум поглощения при 450 нм. Цитохром Р450 содержит протогем, который подобен цитохромам группы b, и участвует в обезвреживании гидрофобных чужеродных для организма молекул (ксенобиотиков). Он является терминальным компонентом микросомальной оксигеназной цепи, обеспечивающей окисление ксенобиотиков. Отдельные разновидности этого цитохрома задействованы в синтезе холестерина, стероидных гормонов и ненасыщенных высших жирных кислот.
В хлоропластах растений содержится представитель цитохромов - цитохром f. Он имеет исключительно растительное происхождение и играет важную роль в переносе электронов по електронотранспортному цепи фотосистемы II хлоропластов.
Дата добавления: 2016-03-05; просмотров: 3620;