Образование билирубина и его метаболизм
Источники билирубина в норме (рис. 38-2). Большинство молекул билирубина образуется в результате катаболизма гемоглобина в стареющих эритроцитах. В норме при этом образуется около 80—85% билирубина в сутки. Когда циркулирующий в крови эритроцит достигает окончания нормального срока своего существования, составляющего приблизительно 120 сут, он разрушается в ретикулоэндотелиальной системе. В процессе катаболизма гемоглобина вначале от гема отделяется глобин, после чего небелковая часть молекулы (ферропротопорфирин IX) разрушается в процессе окисления и превращается в биливердин под действием микросомальной гемоксигеназы. Эта ферментная система нуждается в кислороде и коферменте, представляющим собой восстановленный никотинамиддинуклеотидфосфат (NADPH). Под действием биливердинредуктазы из биливердина затем образуется билирубин (в химической форме билирубина IXa).
Около 15-20% билирубина образуется из других источников. Одним из них служит разрушение созревающих клеток эритроидного ряда в костном мозге, или так называемый неэффективный эритропоэз (см. главу 287). Еще одним источником бывают неэритроидные компоненты (особенно в печени), включающие в себя продукты преобразования гема и близкие к нему по структуре белки (цитохромы, миоглобин и гемсодержащие ферменты). Эти два источника образования билирубина обозначаются совместно как ранняя меченая фракция (термин появился при проведении экспериментов с меченым глицином и D-аминолевулиновой ки слотой — АЛК). Так, когда меченый глицин вводят здоровому человеку, то приблизительно 15% метки обнаруживают в уробилиногенах кала в течение первых 3-5 сут; 85% метки появляется в широком временном диапазоне, пик се определяется приблизительно через 120 сут; эта фракция отражает образование билирубина в норме в результате деструкции стареющих эритроцитов.
Транспорт билирубина. Вслед за высвобождением неконъюгированного билирубина в плазму он фактически весь прочно связывается с альбумином. Максимальная связывающая способность составляет две молекулы билирубина на одну молекулу альбумина, причем связывание происходит обратимым, нековалентным способом. Определенные органические анионы, такие как сульфонамиды и салицилаты, конкурируют с билирубином за общие места связывания на поверхности альбумина и могут вытеснять из них билирубин, что дает ему возможность проникнуть в ткани, например в центральную нервную систему. Большинство данных относительно связывания с альбумином было получено в экспериментах с использованием непрямого билирубина. Прямой билирубин тоже связывается с альбумином, но как обратимым, так и необратимым способами. Обратимое связывание аналогично нековалентному связыванию непрямого билирубина, но связь прямого билирубина наименее прочная. Второй тип связывания приводит к образованию очень прочного, необратимого комплекса альбумин — билирубин, который появляется в сыворотке при нарушении выведения билирубина печенью (например, при холестазе). Вследствие подобного связывания этот комплекс не появляется в моче. Комплекс альбумин — билирубин может также обнаруживаться в сыворотке в течение нескольких недель после устранения обструкции или в период выздоровления больного от печеночной желтухи.
Билирубин обнаруживают в жидкостях организма (в спинномозговой, суставном выпоте, содержимом кист и др.) в пропорциях, соответствующих количеству в них альбумина, но он отсутствует в подлинных секретах, таких как слезы, слюна и сок поджелудочной железы. На развитие желтухи влияют и такие факторы, как кровоток и отеки. Парализованные конечности и отечные участки кожи имеют тенденцию оставаться неокрашенными, при желтухе у больных с гемиплегией и отеками желтушность может быть односторонней.
Метаболизм билирубина в печени (рис. 38-3). Печень играет основную роль в метаболизме желчных пигментов. Различают три фазы метаболизма: 1) поглощение печенью; 2) конъюгация; 3) выведение в желчь. Из этих трех фаз выведение, по-видимому, представляет собой фазу, ограничивающую скорость метаболизма, самую чувствительную к повреждению клеток печени.
Поглощение. Непрямой билирубин, связанный с альбумином, поступает в печеночную клетку, при этом происходит диссоциация пигмента и альбумина. Фаза поглощения и последующего накопления билирубина в гепатоците включает в себя связывание билирубина с определенными цитоплазматическими анион-связанными белками, в частности, с лигандином. Это связывание может предотвратить обратный выход билирубина в плазму.
Связывание. Непрямой билирубин растворим в воде, и, для того чтобы иметь возможность выделяться клетками печени в желчь, он должен превратиться в водорастворимые производные. Это завершается связыванием, вследствие чего большая часть билирубина превращается в глюкуронид билирубина. Эта реакция происходит в эндоплазматической сети гепатоцитов в результате воздействия билирубинглюкуронилтрансферазы. Как показано на рис. 38-3, эта реакция, по-видимому, двухступенчата и приводит вначале к образованию моцоглюкуронида, а затем образуется диглюкуронид. В норме желчь содержит 85% билирубиндиглюкуронида и 15% билирубинмоноглюкуронида. Непрямой билирубин обычно не поступает в желчь (за исключением случаев его фотоокисления, см. ниже).
Экскреция, или секреция, в желчь. В норме для того, чтобы билирубин поступил в желчь, он должен находиться в связанном виде. Несмотря на то что этот процесс в целом недостаточно ясен, секреция прямого билирубина в желчь, по-видимому, представляет собой энергозависямую и ограничивающую скорость метаболизма фазу в метаболизме билирубика в печени. Если она нарушается, то: 1) снижается секреция билирубина в желчь и 2) происходит рсгургитация, или обратный выход прямого билирубина из клеток печени в кровоток.
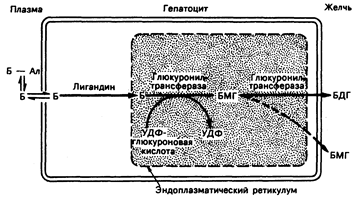
Рис. 38.3. Поглощение билирубина (Б) печенью, его связывания и экскреции в желчь печеночной клеткой.
Несмотря на то что превращение билирубинмоноглюкуронида (БМГ) в билирубин-диглюкуронид (БДГ), по-видимому, катализируется глюкуронилтрансферазой, некоторые исследователи считают обязательным участие в этом превращении мембранной трансглюкуронидазы плазмы. УДФ — уридиндифосфат; Ал -- альбумин.
Как уже было показано, билирубин 1Ха может существовать в виде четырех геометрических изомеров. В норме встречается изомер Z—Z (см. рис. 38-2), т. е. форма, позволяющая связываться водороду внутри молекулы, в результате чего она становится гидрофобной. Другие изомеры (Z—e, e—z и Е—Е, зависящие от положения атомов водорода в двух мостиках двойной связи) могут быть образованы под воздействием синего света и нестабильны. Они водорастворимы, потому что их геометрическая конфигурация препятствует внутримолекулярному связыванию водорода. Таким образом, эти изомеры (фотоизомеры) могут поступать в желчь в несвязанном состоянии. Естественный изомер Z—Z также становится водорастворимым в результате связывания с глюкуроновой кислотой. Образование билирубинглюкуронида предотвращает связывание водорода внутри молекулы, которая приобретает полярность и обеспечивает возможность поступления пигмента в желчь (см. рис. 38-2).
Кишечная фаза метаболизма билирубина. После появления в просвете кишки билирубина глюкуронид может экскретироваться в кал или метаболизироваться до уробилиногена и родственных ему веществ. Вследствие своей полярности прямой билирубин не реабсорбируется слизистой оболочкой кишечника, что представляет собой механизм, способствующий освобождению организма от этого пигмента. Для образования уробилиногена из прямого билирубина необходимо воздействие бактерий, которое происходит в нижнем отделе тонкой кишки и в толстой кишке.
В противоположность прямому билирубину, уробилиноген реабсорбируется из тонкой кишки в портальный кровоток и таким образом становится объектом энтерогепатического круговорота. Часть уробилиногена реэкскретируется печенью в желчь; остальная его часть поступает в мочу в количестве, обычно не превышающем 4 мг/сут. При нарушении механизма печеночной экскреции (например, при гепатоцеллюлярном заболевании) или же значительно усиленном образовании билирубина (например, при гемолитической анемии) концентрация уробилиногена в моче может значительно увеличиться.
Количество выводимого с калом уробилиногена в норме колеблется в пределах 50—280 мг/сут. В условиях сниженной экскреции прямого билирубина в кишечник (например, при болезнях печени, обструкции желчного протока) или подавления микрофлоры кишечника антибиотиками выделение уробилиногена с калом уменьшается. При гемолитической анемии выведение уробилиногена с мочой и калом резко увеличивается.
У здорового человека, у которого объем крови составляет 5 л, а концентрация гемоглобина 150 г/л, общее количество гемоглобина в циркулирующей крови составляет 750 г. Поскольку ежесуточно разрушается приблизительно 0,8% всех эритроцитов, то для катаболизма высвобождается 6,3 г гемоглобина.
Дата добавления: 2016-03-05; просмотров: 1764;