Структура пищевой цепи в морской экосистеме
(по Ф. Рамаду, 1981)
| Трофический уровень | Экологическая функция | Тип организма | Вид | |
| I II III IV V | Продуцент .¯ Консумент 1 (травоядное) ¯ Консумент 11 (плотоядное 1) . ¯ Консумент III (плотоядное 2) ¯ Консумент IV (плотоядное 3) | Фитопланктон ¯ Зоопланктон ¯ Рыбы (микрофаги) ¯ Рыбы (макрофаги) ¯ Птицы (ихтиофаги) | Chaetoceros ¯ Calanus (веслоногие рачки) ¯ Ammodytes (песчанка) ¯ Clupea (сельдь) ¯ Phalacrocorax (баклан) | |
Пищевые цепи, включающие паразитов, отличаются от приведенных и идут от крупных организмов к мелким. В отдельных случаях организмы, таксономически значительно удаленные друг от друга, развиваются один внутри тела другого, первый паразит внутри второго и т. д. К примеру, у насекомых гиперпаразитизм очень сильно развит, и нередко пищевая цепь имеет следующий вид:
I II III IV
растение ® травоядное ® паразит ® гиперпаразит
сосна ® гусеница ® бракониды ® наездники
Abies Choristoneura Apantelessp Chaleididae
alba murinana
Вокруг каждого вида насекомого-фитофага, который питается растениями, формируется зооценоз паразитов и хищников, образующих многочисленные пищевые цепи, где хозяин является начальным звеном.
Приведенные типы пищевых цепей начинаются с фотосинте-зирующих организмов и носят название пастбищных (или цепи выедания, или цепи потребления).
Третий тип пищевых цепей, начинающихся с отмерших остатков растений, трупов и экскрементов животных, относят к детритным (сапрофитным) пищевым цепям или к детрит-ным цепям разложения. В детритных пищевых цепях наземных экосистем важную роль играют лиственные леса, большая часть листвы которых не употребляется в пищу травоядными животными и входит в состав подстилки из опавших листьев. Листья измельчаются многочисленными детритофагами — грибами, бактериями, насекомыми (например, коллембола) и т. д., дальше заглатываются земляными (дождевыми) червями, которые осуществляют равномерное распределение гумуса в поверхностном слое земли, образуя так называемый мулль (рис. 12.22).
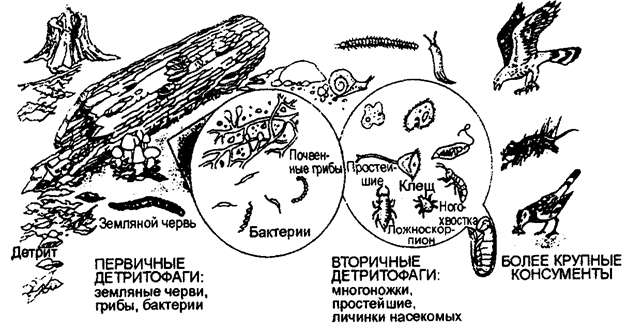
Рис. 12.22. Детритная пищевая цепь в наземной экосистеме
(по Б. Небелу, 1993)
На этом уровне у грибов закладывается мицелий. Разлагающие микроорганизмы, завершающие цепь, производят окончательную минерализацию мертвых органических остатков. В целом типичные детритные пищевые цепи наших лесов можно представить следующим образом:
листовая подстилка ® дождевой червь ® черный дрозд ®
ястреб-перепелятник;
мертвое животное ® личинки падальных мух ® травяная
лягушка ® обыкновенный уж.
В рассмотренных схемах пищевых цепей каждый организм представлен как питающийся другими организмами какого-то одного типа. Реальные же пищевые связи в экосистеме намного сложнее, так как животное может питаться организмами разных типов из одной и той же пищевой цепи или из разных пищевых цепей, например, хищники верхних трофических уровней. Нередко животные питаются как растениями, так и другими животными. Их называют всеядными. Таким образом, все три типа пищевых цепей всегда сосуществуют в экосистеме так, что ее представители объединены многочисленными пересекающимися пищевыми связями, а все вместе они образуют пищевую (трофическую) сеть (рис. 12.23).
Пищевые сети в экосистемах весьма сложные, и можно сделать вывод, что энергия, поступающая в них, долго мигрирует от одного организма к другому.
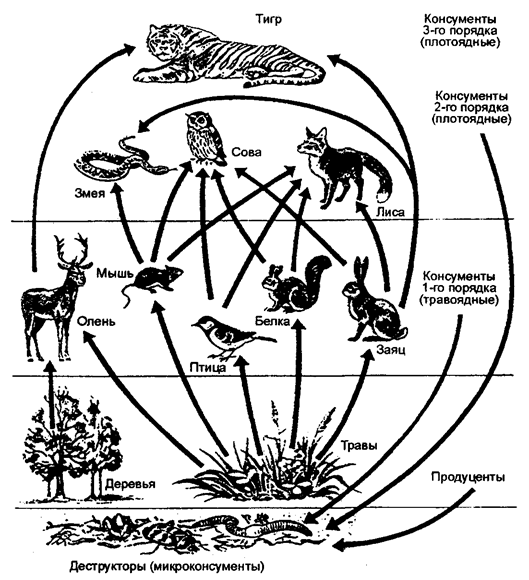
Рис. 12.23. Пищевая сеть и направление потока вещества
(по Е. А. Криксунову и др., 1995)
Экологические пирамиды. Внутри каждой экосистемы трофические сети имеют хорошо выраженную структуру, которая характеризуется природой и количеством организмов, представленных на каждом уровне различных пищевых цепей. Для изучения взаимоотношений между организмами в экосистеме и для их графического изображения обычно используют не схемы пищевых сетей, а экологические пирамиды. Экологические пирамиды выражают трофическую структуру экосистемы в геометрической форме. Они строятся в виде прямоугольников одинаковой ширины, но длина прямоугольников должна быть пропорциональна значению измеряемого объекта. Отсюда можно получить пирамиды численности, биомассы и энергии.
Экологические пирамиды отражают фундаментальные характеристики любого биоценоза, когда они показывают его трофическую структуру:
— их высота пропорциональна длине рассматриваемой пищевой цепи, т. е. числу содержащихся в ней трофических уровней;
— их форма более или менее отражает эффективность превращений энергии при переходе с одного уровня на другой.
Пирамиды численности. Они представляют собой наиболее простое приближение к изучению трофической структуры экосистемы. При этом сначала подсчитывают число организмов на данной территории, сгруппировав их по трофическим уровням и представив в виде прямоугольника, длина (или площадь) которого пропорциональна числу организмов, обитающих на данной площади (или в данном объеме, если это водная экосистема). Установлено основное правило, которое гласит, что в любой среде растений больше, чем животных, травоядных больше, чем плотоядных, насекомых больше, чем птиц, и т. д. (рис. 12.24).

Рис. 12.24. Упрощенная схема пирамиды численности
(по Г. А. Новикову, 1979)
Пирамиды численности отражают плотность организмов на каждом трофическом уровне. В построении различных пирамид численности отмечается большое разнообразие. Нередко они перевернуты (рис. 12.25).
Например, в лесу насчитывается значительно меньше деревьев (первичные продуценты), чем насекомых (растительноядные).
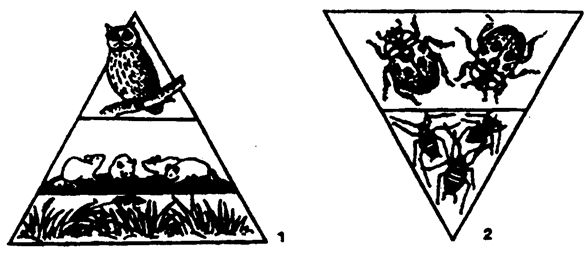
Рис. 12.25. Пирамиды численности:
1 — прямая; 2 — перевернутая (по Е. А. Криксунову и др., 1995)
Подобная же картина наблюдается в пищевых цепях сапрофитов и паразитов.
Пирамида биомассы. Отражает более полно пищевые взаимоотношения в экосистеме, так как в ней учитывается суммарная масса организмов (биомасса) каждого трофического уровня. Прямоугольники в пирамидах биомассы отображают массу организмов каждого трофического уровня, отнесенную к единице площади или объема. Форма пирамиды биомассы нередко сходна с формой пирамиды численности. Характерно уменьшение биомассы на каждом следующем трофическом уровне (рис. 12.26 и 12.27).


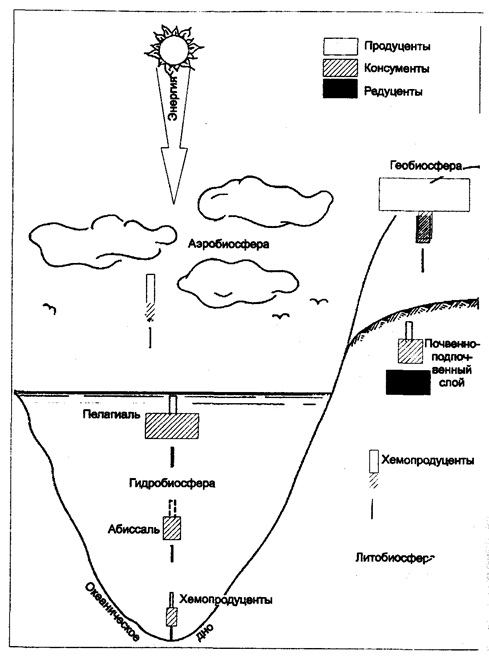
Рис. 12.27. Типы пирамид биомассы в различных подразделениях
биосферы (по Н. Ф. Реймерсу, 1990)
Пирамиды биомассы, так же как и численности, могут быть не только прямыми, но и перевернутыми. Перевернутые пирамиды биомассы свойственны водным экосистемам, в которых первичные продуценты, например фитопланктонные водоросли, очень быстро делятся, а их потребители — зоопланктонные ракообразные — гораздо крупнее, но имеют длительный цикл воспроизводства. В частности, это относится к пресноводной среде, где первичная продуктивность обеспечивается микроскопическими организмами, скорость обмена веществ которых повышена, т. е. биомасса мала, производительность велика.
Пирамида энергии. Наиболее фундаментальным способом отображения связей между организмами наразных трофических уровнях служат пирамиды энергии. Они представляют эффективность преобразования энергии и продуктивность пищевых цепей, строятся подсчетом количества энергии (ккал), аккумулированной единицей поверхности за единицу времени и используемой организмами на каждом трофическом уровне. Так, можно относительно легко определить количество энергии, накопленной в биомассе, и сложнее оценить общее количество энергии, поглощенной на каждом трофическом уровне. Построив график (рис. 12.28), можно констатировать, что деструкторы, значимость которых представляется небольшой в пирамиде биомассы, а в пирамиде численности наоборот; получают значительную часть энергии, проходящей через экосистему. При этом только часть всей этой энергии остается в организмах на каждом трофическом уровне экосистемы и сохраняется в биомассе, остальная часть используется для удовлетворения метаболических потребностей живых существ: поддержание существования, рост, воспроизводство. Животные также расходуют значительное количество энергии и для мышечной работы.

Рис. 12.28. Экологические пирамиды (по Е. Одуму, 1959):
а — пирамида численности; б — пирамида биомассы;
в — пирамида энергии.
Заштрихованные прямоугольники обозначают чистую продукцию
Рассмотрим более подробно, что происходит с энергией при ее передаче через пищевую цепь (рис. 12.29).
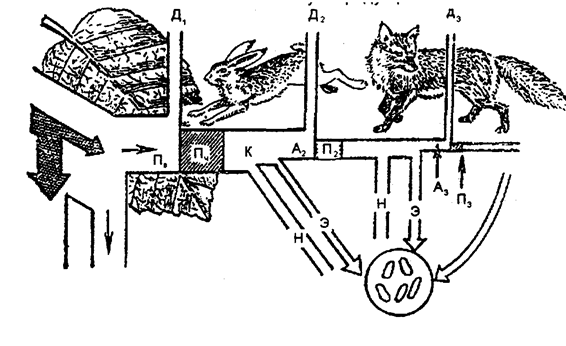
Рис. 12.29. Поток энергии через три уровня трофической
цепи (по П. Дювиньо и М. Тангу, 1968)
Ранее уже было отмечено, что солнечная энергия, полученная растением, лишь частично используется в процессе фотосинтеза. Фиксированная в углеводах энергия представляет собой валовую продукцию экосистемы (Пв). Углеводы идут на построение протоплазмы и рост растений. Часть их энергии затрачивается на дыхание (Д1). Чистая продукция (Пч) определяется по формуле:
Пч = Пв – Д1 (12.5)
Следовательно, поток энергии, проходящий через уровень продуцентов, или валовую продукцию, можно представить:
Пв = Пч + Д1. (12.6)
Определенное количество созданных продуцентами веществ служит кормом (К) фитофагов. Остальное как итог отмирает и перерабатывается редуцентами (Н). Ассимилированный фитофагами корм (А) лишь частично используется для образования их биомассы (Пд). Главным образом он растрачивается на обеспечение энергией процессов дыхания (Д) и в определенной степени выводится из организма в виде выделений и экскрементов (Э). Поток энергии, проходящий через второй трофический уровень, выражается следующим образом:
А2 =П2 + Д2. (12.7)
Консументы второго порядка (хищники) не истребляют всю биомассу своих жертв. При этом из того количества ее, которое они уничтожают, только часть используется на создание биомассы их собственного трофического уровня. Остальная же часть в основном затрачивается на энергию дыхания, выделяется с экскретами и экскрементами. Поток энергии, проходящий через уровень консументов второго порядка (плотоядные), выражается формулой:
А3 = П3 + Д3. (12.8)
Подобным образом можно проследить совокупность пищевой цепи и до последнего трофического уровня. Распределив по вертикали различные затраты энергии на трофических уровнях, получим полную картину пищевой пирамиды в экосистеме (рис. 12.30).

Рис. 12.30. Пирамида энергии (из Ф. Рамада, 1981):
Е - энергия, выделяемая с метаболитами; D - естественные смерти; W —фекалии; R - дыхание
Поток энергии, выражающийся количеством ассимилированного вещества по цепи питания, на каждом трофическом уровне уменьшается или:
Пч > П2 > П3 и т.д.
Р. Линдеман в 1942 г. впервые сформулировал закон пирамиды энергий, который в учебниках нередко называют «законом 10%». Согласно этому закону с одного трофичес-когоуровня экологической пирамиды переходит на другой ее уровень в среднем не более 10% энергии.
Последующим гетеротрофам передается только 10—20% исходной энергии. Используя закон пирамиды энергий, нетрудно подсчитать, что количество энергии, доходящее до третичных плотоядных (V трофический уровень), составляет около 0,0001 энергии, поглощенной продуцентами. Отсюда следует, что передача энергии с одного уровня на другой происходит с очень малым КПД. Это объясняет ограниченное количество звеньев в пищевой цепи независимо от того или иного биоценоза.
Е. Одум (1959) в предельно упрощенной пищевой цепи - люцерна ® теленок ® ребенок оценил превращение энергии, проиллюстрировал величину ее потерь. Допустим, рассуждал он, имеется посев люцерны на площади 4 га. На этом поле кормятся телята (предполагается, что они едят только люцерну), а 12-летний мальчик питается исключительно телятиной. Результаты расчетов, представленные в виде трех пирамид: численности, биомассы и энергии (рис. 12.31 и 12.32), — свидетельствуют; что люцерна использует всего 0,24% всей падающей на поле солнечной энергии, теленком усваивается 8% этой продукции и только 0,7% биомассы теленка обеспечивает развитие ребенка в течение года*.
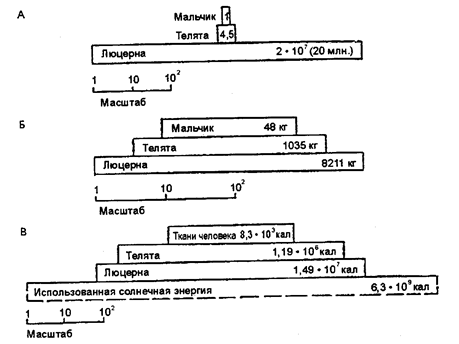
Рис. 12.31. Упрощенная экосистема: люцерна — телята — мальчик
(по Е. Одуму, 1959):
А — пирамида чисел; Б — пирамида биомассы; В — пирамида энергии

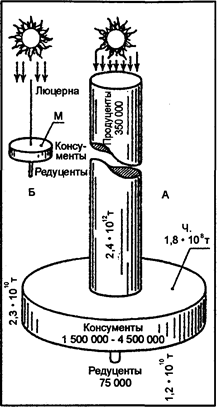
Е. Одум, таким образом, показал, что только одна миллионная доля приходящейся солнечной энергии превращается в биомассу плотоядного, в данном случае способствует увеличению массы ребенка, а остальное теряется, рассеивается в деградированной форме в окружающей среде. Приведенный пример наглядно иллюстрирует очень низкую экологическую эффективность экосистем и малый КПД при превращении в пищевых цепях. Можно констатировать следующее: если 1000 ккал (сут м2) зафиксирована продуцентами, то 10 ккал (сут. м2) переходит в биомассу травоядных и только 1 ккал (сут. м2) — в биомассу плотоядных.
Поскольку определенное количество вещества может быть использовано каждым биоценозом неоднократно, а порция энергии один раз, то целесообразнее говорить, что в экосистеме происходит каскадный перенос энергии (см. рис. 12.19).
Консументы служат управляющим и стабилизирующим звеном в экосистеме (рис. 12.32). Консументы порождают спектр разнообразия в ценозе, препятствуя монополии доминантов. Правило управляющего значения консументов можно с полным основанием отнести к достаточно фундаментальным. Согласно кибернетическим воззрениям, управляющая система должна быть сложнее по структуре, чем управляемая, то становится ясной причина множественности видов консументов. Управляющее значение консументов имеет и энергетическую подоснову. Поток энергии, проходящий через тот или другой трофический уровень, не может абсолютно определяться наличием пищи в нижележащем трофическом уровне. Всегда остается, как известно, достаточный «запас», так как полное уничтожение корма привело бы к гибели потребителей. Эти общие закономерности наблюдаются в рамках популяционных процессов, сообществ, уровней экологической пирамиды, биоценозов в целом.
Дата добавления: 2016-02-24; просмотров: 742;