Понятие о популяции в экологии
Любой вид животного, растения, микроорганизма утверждает себя во внешней среде, приспосабливается к постоянно изменяющимся условиям не как простая сумма особей, а в форме своеобразных группировок организмов, представляющих единое функциональное целое — популяцию (от лат. populus — народ, население).
Каждый вид занимает тот или иной ареал, в различных частях которого наблюдаются разные условия, и чем больше ареал, тем существеннее эти различия. Часто группировки особей, располагающиеся в разных частях ареала вида, настолько изолированы друг от друга, что не могут вступать в контакты и не скрещиваются. Количество группировок зависит от численности и исторического (филогенетического) возраста вида, размеров ареала и от других причин. Группировки особей с общим генофондом, сходной морфологией, единым жизненным циклом и представляют собой популяции.
По С. С. Шварцу, популяция — эго элементарная группировка организмов определенного вида, обладающая всеми необходимыми условиями для поддержания своей численности необозримо длительное время в постоянно изменяющихся условиях среды. Это свидетельствует о том, что приспособительные возможности популяции гораздо выше, чем у слагающих ее индивидов.
Популяция — это совокупность особей одного вида, которая обладает общим генофондом и занимает определенную территорию. В современной экологии популяция рассматривается как элементарная единица процесса микроэволюции, которая способна реагировать на изменения среды перестройкой своего генофонда.
Поскольку все виды живых организмов представлены в природе конкретными популяциями, понятие «популяция» является одним из центральных в биологии, а генетические, экологические, эволюционные подходы к изучению популяций объединяются в особое направление — популяционную биологию.
Таким образом, вид— это сложная биологическая система, состоящая из группировок организмов — популяций, обладающих характерными особенностями строения, физиологии и поведения. Такие внутривидовые группировки являются генетической единицей вида, а изменения их обусловливают эволюцию данного вида, т. е. популяции служат элементарными эволюционирующими структурами. Следовательно, популяция как группа совместно обитающих особей одного вида выступает первой надорганизменной биологической макросистемой.
Популяция как биологическая единица обладает определенной структурой и функцией. Структура популяции характеризуется составляющими ее особями и их распределением в пространстве. Функции популяции тождественны функциям других биологических систем. Популяциям свойствен рост, развитие, способность поддерживать существование в постоянно меняющихся условиях, т. е. популяция обладает конкретными генетическими и экологическими характеристиками.
Широкое распространение в экологии получила концепция иерархии популяций в зависимости от размеров занимаемой ими территории. Н. П. Наумов, например, различает элементарную, экологическую и географическую популяции :
- элементарная (локальная) популяция — это совокупность особей вида, занимающих какой-то небольшой участок однородной площади. Количество элементарных популяций, на которые распадается вид, зависит от разнородности условий в биогеоценозе: чем они однообразнее, тем меньше элементарных популяций, и наоборот.
Нередко смешение особей элементарных популяций, происходящее в природе, стирает границы между ними.
- экологическая популяция формируется как совокупность элементарных популяций. В основном это внутривидовые группировки, приуроченные к конкретным биогеоценозам. Например, белка (Sciurus vulgaris) заселяет различные типы леса. Поэтому могут быть четко выделены «сосновые», «елово-пихтовые» и другие экологические популяции ее. Они слабо изолированы друг от друга, и обмен генетической информацией между ними происходит довольно часто, но реже, чем между элементарными популяциями.
- географические популяции слагаются из экологических и охватывают группу особей, заселяющих территорию с географически однородными условиями существования. Географические популяции довольно основательно разграничены и относительно изолированы. Они различаются плодовитостью, размерами особей, рядом экологических, физиологических, поведенческих и других особенностей. Примером могут служить популяции белки в заенисейской тайге и смешанных лесах Беларуси, степная и тундровая популяции узкочерепной полевки (Microtus gregalis).
Если под влиянием ряда факторов географическая популяция приобретает устойчивые особенности, отчетливо выделяющие ее из соседних, такая популяция называется географической расой или подвидом. В целом такие понятия, как географическая популяция, географическая раса, подвид, часто рассматриваются как синонимы. Например, вид «белка обыкновенная» насчитывает более 20 географических популяций, или рас, или подвидов.
В природе границы и размеры популяций определяются особенностями не только заселяемой территории, но и главным образом свойствами самой популяции. В основе всегда лежит степень ее генетического и экологического единства. Как показывает Н. П. Наумов, раздробление вида на множество мелких территориальных группировок является процессом приспособления к огромному разнообразию местных условий. Это увеличивает генетическое многообразие вида и обогащает его генофонд.
Основными характеристиками популяции являются: плотность, численность, рождаемость, смертность, возрастной состав, характер распределения в пределах территории и тип роста.
Плотность популяции определяется числом особей, приходящихся единицу площади или объема. Территория, занимаемая разными популяциями одного вида, а тем более различных видов, очень колеблется и зависит от степени подвижности особей. Так, одна популяция ящерицы прыткой занимает от 0,1 до нескольких гектаров; северного оленя — более 100 км2. Каждому виду присуща определенная оптимальная плотность популяции, отклонение от которой в обе стороны отрицательно сказываются на темпах воспроизводства и жизнедеятельности особей.
Численность или число особей в популяции различна у разных видов но она не может быть ниже некоторых пределов. Сокращение численности за эти пределы может привести к вымиранию популяции. Численность популяций может резко меняться по сезонам и годам. У видов животных и растений с большой продолжительностью жизни и относительно малой плодовитостью численность популяций более устойчивая. Колебания численности относительно оптимального уровня носят разный характер. Они могут б плавными, циклическими и резкими, нерегулярными. При этом быстрое возрастание численности начинается обычно под действием модифицирующих факторов, не зависящих от плотности популяции, и чаще в благоприятных погодных условий, которые действуют на популяцию прямо и косвенно. Различают межвидовые и внутривидовые механизмы регуляции численности. К первым относят паразитов, хищников и болезни, ко вторым — взаимодействия особей внутри популяции. В простейшем виде это разнообразные формы внутривидовой конкуренции, связанные с прямым и косвенным отрицательным воздействием особей друг на друга. Полагают, что при численности популяции меньше нескольких сотен особей случайные причины (пожар, наводнение, изменение погоды) могут сократить ее настолько, что рождаемость перестанет перекрывать убыль и на протяжении нескольких поколений оставшиеся особи вымрут.
Рождаемость популяции определяется прежде всего эволюционным положением вида, его биологией. Низкая плодовитость характерна для техвидов, которые проявляют большую заботу о потомстве. Кроме того, рождаемость зависит от скорости полового созревания, числа генераций в году, соотношения в популяции самцов и самок, обеспеченности кормом, влияния погодных условий.
Смертность популяции — это количество особей, погибших за определенный период. Она бывает очень высокой и изменяется в зависимости от условий среды, возраста и состояния популяции. У большинства видов смертность в раннем возрасте всегда бывает выше, чем у взрослых особей. Однако встречаются и такие виды, у которых смертность приблизительно одинакова во всех возрастах или преобладает у особей старших возрастов. Факторы смертности очень разнообразны. Смертность может быть вызвана влиянием физических условий низкие и высокие температуры, ливневые осадки, град, избыточная и недостаточная влажность и др.), биотическими факторами (отсутствие корма, инфекционные заболевания и т. д.), антропогенными факторами загрязнение окружающей среды, уничтожение животных, вырубка деревьев и др.).
Возрастной состав популяции имеет большое значение для ее существования и процветания. При благоприятных условиях в популяции наличествуют все возрастные группы и поддерживается более или менее стабильный возрастной состав. В быстро растущих популяциях доминируют интенсивно размножающиеся молодые особи, а в сокращающихся — старые, уже неспособные интенсивно размножаться. Такие популяции малопродуктивны, недостаточно устойчивы. Возрастной состав популяции зависит от продолжительности жизни особей, периода достижения ими половой зрелости, числа генераций.
Популяция состоит из разных по полу и возрасту особей. Соотношение половозрелых и неполовозрелых особей в популяциях различно и за висит от продолжительности жизни, времени наступления половой зрелости, интенсивности размножения.
Динамика популяций
Динамика численности и плотности популяций находится в тесной зависимости от плодовитости и смертности, а также от способности слагающих их особей совершать миграции.
В живых организмах заложена огромная возможность к размножению. Скорость увеличения численности особей популяции при отсутствии лимитирующих факторов обозначается как условный показатель — биотический потенциал. Он устанавливается теоретически как возможная средняя величина рождаемости или скорость, с которой при беспрерывном размножении (возможном только теоретически) особи определенного вида могут покрыть земной шар равномерным споем. Для слонов эта скорость составляет 0,3 м/с, а для некоторых микроорганизмов — сотни метров в секунду. Подсчитано, что бактерии делятся каждые 20 минут. При таком темпе одна клетка за 36 часов дает потомство, которое может покрыть сплошным слоем всю нашу планету. Один одуванчик менее чем за 10 лет способен заселить своими потомками земной шар, если все семена прорастут. В действительности же такая громадная плодовитость организмов никогда не реализуется. Истинная плодовитость значительно ниже потенциальной. Это наглядно прослеживается на примере капустной белянки. В результате гибели яиц, гусениц и куколок по разным причинам (болезни, паразиты, хищники и др.) на свет появляется лишь 0,32% взрослых бабочек от числа отложенных яиц.
Средняя величина плодовитости каждого вида определилась исторически как приспособление, обеспечивающее пополнение убыли популяций. Поэтому естественно, что у менее приспособленных к неблагоприятным условиям видов высокая смертность в молодом (личиночном) возрасте компенсируется значительной плодовитостью. Это особенно типично для организмов, подвергающихся интенсивной неизбирательной элиминации. Например, среди насекомых самая высокая плодовитость у растительноядных форм, а наиболее низкая — у паразитов и хищников.
В благоприятных условиях плодовитость обычно низкая. Этим объясняется более высокая плодовитость животных в зонах умеренного климата по сравнению с тропическими видами. Кроме того, характер плодовитости зависит от скорости полового созревания, числа генераций в течение сезона, от соотношения в популяции самцов и самок. Если вид размножается с большой скоростью и чутко реагирует на изменения условий среды, то численность популяций его быстро и существенно изменяется. Примером могут служить многие насекомые и мышевидные грызуны.
Различают абсолютную и удельную рождаемость. Первая характеризуется общим числом родившихся особей. Например, если в популяции северных оленей, насчитывающей 16 тыс. голов, за год появилось 2 тыс. оленят, то это число и выражает абсолютную рождаемость. Удельную рождаемость вычисляют как среднее изменение численности на особь за определенный интервал времени, и в данном случае она составит 0,125, т.е. один новорожденный на восемь членов популяции за год.
Величина рождаемости зависит от многих причин. Большое значение имеет доля особей, способных в данный период к размножению, что определяется соотношением полов и возрастных групп. Важна также частота последовательности генераций.
Для величины рождаемости имеет значение также соотношение периода размножения и общей продолжительности жизни. Этот период для самок дрозофил составляет около 65%, для стадной саранчи шистоцерки — 15%, а для поденок — всего от 0,5 до 1% длительности их существования. Большую роль играет плодовитость особей. Однако размножаемость популяции, как правило, не бывает прямо пропорциональна плодовитости. Плодовитость сильно зависит от степени развития заботы о потомстве или обеспеченности яиц питательными материалами.
У видов, оберегающих и выкармливающих свой молодняк, плодовитость резко понижена. Размер кладки у птиц разных видов варьирует уже не в тысячи раз, как у рыб, а в пределах от одного яйца (у некоторых хищников, пингвинов, и др.) до 20—25 (максимальное количество яиц у куриных, например, у серой куропатки).
Большая плодовитость вырабатывается у видов также в условиях более высокой смертности, особенно при сильном давлении со стороны хищников. Отбор на более высокую плодовитость — одни из приспособлений, компенсирующих большую норму гибели популяциях. Поэтому при высокой плодовитости общая размножаемость популяции может быть весьма низкой. В различных популяциях одного вида плодовитость обычно тем выше, чем более неблагоприятны занимаемые ими места обитания. Так, у многих млекопитающих — зайцев, мышей, полевок — число детенышей в помете на границах ареала больше, чем в его центре.
Численность и плотность популяции зависят также от ее смертности. Смертность популяции— это количество особей, погибших за определенный период. - Она, как и плодовитость, изменяется в зависимости от условий среды, возраста и состояния популяции и выражается в процентах к начальной или чаще к средней величине ее. У большинства видов смертность в раннем возрасте всегда выше, чем у взрослых особей. У многих рыб до взрослой фазы доживает 1—2% от числа выметанной икры, у насекомых — 0,3—0,5% от отложенных яиц.
Различают три типа смертности. К первому относится смертность, одинаковая во всех возрастах. Она выражается экспоненциальной кривой (убывающей геометрической прогрессией).
Такая смертность встречается очень редко и только у популяций, постоянно находящихся в оптимальных условиях.
Второй тип смертности отличается повышенной гибелью особей на ранних стадиях развития и свойствен большинству растений и животных. Максимальная гибель многих растений происходит в стадии прорастания семян и всходов, а животных — в личиночной фазе или в молодом возрасте.
Третий тип смертности характеризуется повышенной гибелью взрослых, главным образом старых особей. Наблюдается он у насекомых, личинки которых обитают в почве, воде, древесине или в других местах с благоприятными условиями, а также у проходных рыб, которые нерестуют один раз в жизни.
Обычно смертность представляется графически. Широкое распространение получило построение «кривых выживания». Они выражают зависимость числа выживших из 100 или 1000 особей от их возраста. Соответственно трем типам смертности было получено три типа кривых (рис. 29).
На основании данных, получаемых при исследовании закономерностей плодовитости и смертности популяций, появляется возможность математического моделирования динамики популяций, что имеет огромное теоретическое и практическое значение.
Например, если учитывать, что лишь немногие организмы в течение жизни реализуют свой репродуктивный материал (потенциальные возможности размножения), практический интерес представляет определение так называемой чистой скорости размножения r0. Под r0 подразумевается среднее число потомков, производимых одной особью данного вида за всю жизнь. Если обозначить через х возраст особи, через lХ — выживаемость, т. е. долю особей, доживших до возраста х, через тх — плодовитость, т. е. число потомков, произведенное особью в возрасте х, то произведение lхтх дает ожидаемое число потомков. Суммируя это произведение по всем возрастам от 0 до ∞ при дискретной градации возрастных классов для rq, получим
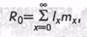 ,
,
а при непрерывной градации классов,
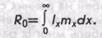 .
.
В реальном случае вместо нижнего предела интегрирования следует брать а — возраст первого размножения, а вместо верхнего предела со — возраст последнего размножения. Тогда второе выражение приобретает следующий вид:
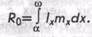
При этом важно иметь в виду, что для любого возраста, в котором не происходит размножение, произведение 1хтх равно 0.
На динамику численности популяции большое влияние оказывают миграции отдельных особей, т. е. обмен индивидуумов популяций. Это происходит при переселении их из одного местообитания в другое. Вместе с плодовитостью и смертностью миграции определяют характер роста популяции и ее плотность. В основном миграции совершаются при подрастании молодняка и его расселении. Наиболее интенсивно расселение происходит при несбалансированных рождаемости и смертности. В результате миграций устраняется избыток особей в одной популяции и компенсируется недостаток в другой, в которой по каким-то причинам смертность превышает рождаемость. В разных популяциях рождаемость, смертность и миграции различаются. Это приводит к сезонным и многолетним колебаниям численности популяции. Они специфичны для каждого вида и зависят от множества причин, обусловливающих особенности плодовитости, смертности и подвижности особей.
Популяция регулирует свою численность и приспосабливается к изменяющимся условиям среды путем обновления и замещения особей. Особи появляются в популяции благодаря рождению и иммиграции, а исчезают в результате смерти и эмиграции. При сбалансированной интенсивности рождаемости и смертности формируется стабильная популяция. В такой популяции смертность компенсируется приростом и численность ее, а также ареал удерживаются на одном уровне. Однако любая популяция наделена как статическими, так и динамическими свойствами и ее плотность постоянно изменяется. Но при более или менее стабильных внешних условиях и состоянии самой популяции колебания эти происходят около какой-то средней величины. В результате популяции не сокращают и не увеличивают численности, не расширяют и не сужают своего ареала. В таком случае они должны в каждом поколении воспроизводить по паре особей от каждой пары. Если это действительно наблюдается, можно говорить о стабильной популяции. Фактически же в природе не известно ни одной популяции, которая сохранялась бы неизменной хотя бы на протяжении короткого промежутка времени.
Чаще отмечается превышение рождаемости над смертностью, и численность популяции растет вплоть до такой степени, что может наступить вспышка массового размножения. Это особенно характерно для мелких животных. Подобные популяции будут растущими. Примером может служить интенсивное размножение и распространение ондатры (Ondatra zibetica) в Европе, начавшееся в 1905 г. в окрестностях Праги, когда там было выпущено 5 особей. Увеличение численности популяции характерно для колорадского жука (Leptinotarsa decemlineata), быстро расселившегося из Франции до Украины, Беларуси и даже до Смоленской и Псковской областей. За последние десятилетия наметился рост популяции канареечного вьюрка (Serinus canaria), зеленой пеночки (Phylloscopus trochiloides), чайки обыкновенной, зайца-русака и некоторых других видов, продвигающихся на северо-восток европейской части. В прудах Подмосковья отмечается столь значительный рост численности ротана (Perccottus glehni), попавшего сюда случайно, что он вытесняет остальных рыб.
Аналогичные примеры растущих популяций известны и для растений. Таковы аистник (Erodium) и горчица (Brassica), завезенные в Калифорнию, а также элодея, появившаяся в 1836 г. в Ирландии и уже в 1885 г. проникшая в бассейн Оки.
Однако при чрезмерном развитии популяции ухудшаются условия существования, что вызывается ее переуплотнением. Это приводит к резкому возрастанию смертности, и в результате численность популяции начинает сокращаться. Так произошло со всеми популяциями элодеи в названных водоемах, ондатры в Беларуси и на Украине, домового воробья (Passer domesticus) в умеренной зоне. Если смертность превышает рождаемость, популяция становится сокращающейся.
Хорошо известны случаи резкого сокращения численности популяций промысловых видов животных, усиленно уничтожаемых человеком (соболь — Martes zibellina, бобр — Castor fiber, выхухоль — Desmana moschata, зубр — Bison bonasus и др.). Распашка степей привела к сокращению численности и ареалов популяций стрепета (Otis tetrax) и дрофы (О. tarda). Реликтовая фауна — также пример сокращающихся в связи с изменением среды популяций. В настоящее время таких популяций довольно мапо и они немногочисленны. Это популяции султанской курицы (Parphirio palyocephalus), кулана (Equus hemionus), окапи (Okapia johnstoni), растений гинкго (Ginkgo biloba) и индийского лотоса (Nelumba nucibera).
Однако сокращаться безгранично популяция также не может. При определенном уровне численности интенсивность смертности начинает падать, а плодовитости — повышаться. В итоге сокращающаяся популяция, достигнув какой-то минимальной численности, превращается в свою противоположность — в растущую популяцию. Рождаемость в такой популяции постепенно растет и в определенный момент выравнивается со смертностью, т. е. популяция на короткий промежуток времени становится стабильной, а затем растущей.
Обычно различают два типа роста популяции: экспоненциальный (логарифмический) и логистический. Экспоненциальный рост описывается уравнением
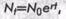
где Nf — численность популяции в момент времени t; N0 — численность популяции в начальный момент t0; е — основание натуральных логарифмов (2,7182); r— показатель темпов размножения. Это уравнение выражает биотический потенциал.
Для того чтобы экспоненциальный рост численности продолжался в течение некоторого времени, необходимо, чтобы значение г было постоянным. Поэтому экспоненциальный рост возможен только в стабильных условиях при отсутствии ограничивающих факторов и в природе либо не происходит вообще, либо происходит в течение очень непродолжительного времени.
Математические формулы экспоненциального роста были приведены А. Лоткой в 1920-х гг., но были известны задолго до этого. В частности, способность популяции расти по экспоненциальному закону явилась одной из основных предпосылок теории естественного отбора. В настоящее время уравнения, описывающие экспоненциальный рост, используются в экологии прежде всего для выяснения потенциальных возможностей популяций к росту.
В связи с тем что рост популяции по экспоненциальному закону продолжается очень короткое время и по мере увеличения численности скорость ее роста снижается, часто описание этого процесса выражается уравнением логистического роста. В основе логистического уравнения заложено предположение, что снижение скорости удельного роста
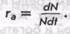
По мере возрастания численности N эта скорость при достижении предельной для данной среды численности К становится равной 0. Следовательно, если N=K, то га=0. Логистическое уравнение нагляднее всего записывается в дифференциальной форме
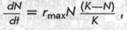
где rmax — константа экспоненциального роста, который мог бы наблюдаться при N=Q, т. е. в начальный момент увеличения численности.
В связи с тем что такой рост популяций наблюдается очень часто, логистическое уравнение, которое описывает его, позволяет судить об общих закономерностях этого процесса. Константы r и К из логистического уравнения дали название двум типам естественного отбора.
Периодические и непериодические колебания численности популяций под влиянием абиотических и биотических факторов среды, свойственные всем видовым популяциям, называются популяционными волнами.
Исходя из предложенных выше расчетов, это можно выразить следующим образом: если r0>], популяция растет, если r0=1, популяция стабильна, а если r0<1, — сокращается.
Дата добавления: 2016-02-09; просмотров: 4525;