Глава 21. Обмен сложных липидов
К сложным липидам относят такие соединения, которые, помимо липидного, содержат и нелипидный компонент (белок, углевод или фосфат). Соответственно существуют протеолипиды, гликолипиды и фосфолипиды. В отличие от простых липидов, используемых в качестве энергетического материала, сложные липиды выполняют пластические функции и используются главным образом как структурные компоненты биологических мембран. Протеолипиды являются структурными компонентами в миелиновых оболочках нервных клеток, в синаптических мембранах и внутренних мембранах митохондрий. Гликолипиды участвуют в функционировании мембран: вовлечены в процессы рецепции, участвуют в контроле и регуляции межклеточных контактов. Обладают высокой тканевой специфичностью и выступают в роли антигенов клеточной поверхности. Фосфолипиды (ФЛ) играют важную роль в структуре и функционировании клеточных мембран, активации мембранных и лизосомальных ферментов, в проведении нервных импульсов, свертывании крови, иммунологических реакциях, процессах клеточной пролиферации и регенерации тканей, в переносе электронов в ЦТД.
Образование ФЛ наиболее интенсивно происходит в печени, стенке кишечника, семенниках, яичниках и молочной железе. Синтез ФЛ, содержащих холин и этаноламин начинается с активации азотистых оснований при участии АТФ и соответствующих киназ. При синтезе фосфатидилинозитола на первом этапе происходит взаимодействие фосфатидной кислоты с ЦТФ, ведущее к образованию цитидиндифосфатдиацилглицерола, который реагирует с инозитолом, образуя фосфатидилинозитол.
Помимо путей синтеза индивидуальных ФЛ, имеются пути их взаимопревращений, целесообразность которых, очевидно, связана с необходимостью обеспечения тканей требуемым ФЛ в нужный момент.
Для синтеза фосфатидилхолинов, и в меньшей степени – сфингомиелинов, нужен холин или метионин, потребность в которых в значительной степени покрывается за счет пищевых источников. При длительном недостатке в пище холина и метионина наблюдается развитие жировой инфильтрации печени, при которой содержание липидов, главным образом ТАГ, может достигать в расчете на сухую массу ткани 45 %, против 7–14 % в норме. Механизм развития жировой инфильтрации печени связан с недостаточным синтезом фосфатидилхолинов и сфингомиелинов, необходимых для формирования в этом органе ЛП. На образование последних, наряду с ФЛ, используются значительные количества ТАГ и холестерола. Сформированные в печени ЛП, в частности богатые триацилглицеролами ЛПОНП, поступают в кровяное русло. Следовательно, образование ЛП можно рассматривать как важнейший путь утилизации печеночных липидов. Поэтому недостаточный синтез в печени содержащих холин ФЛ нарушает образование ЛП и ведет к накоплению в этом органе ТАГ и ХС. По этой причине холин, метионин, а также фосфатидилхолин относятся к группе липотропных веществ, прием которых с пищей предотвращает развитие жировой инфильтрации печени.
Распад фосфолипидов может происходить при участии нескольких ферментов, каждый из которых катализирует гидролитический разрыв строго определеннной связи. Гидролиз некоторых ФЛ под действием фосфолипаз имеет значение не только как путь катаболизма, но и как путь образования эйкозаноидов. Кроме того, фосфолипазы А1 и А2 участвуют в изменении состава жирных кислот в ФЛ, например при синтезе в эмбриональном периоде дипальмитоилфосфатидилхолина – компонента сурфактанта.
Для образования гликолипидов и сфингомиелина (сфинголипидов) вначале требуется синтез самого сфингозина. Это происходит путем конденсации пальмитоил‑КоА с серином при участии пиридоксальфосфата (ПАЛФ) и ионов марганца (Рис. 21.1.).
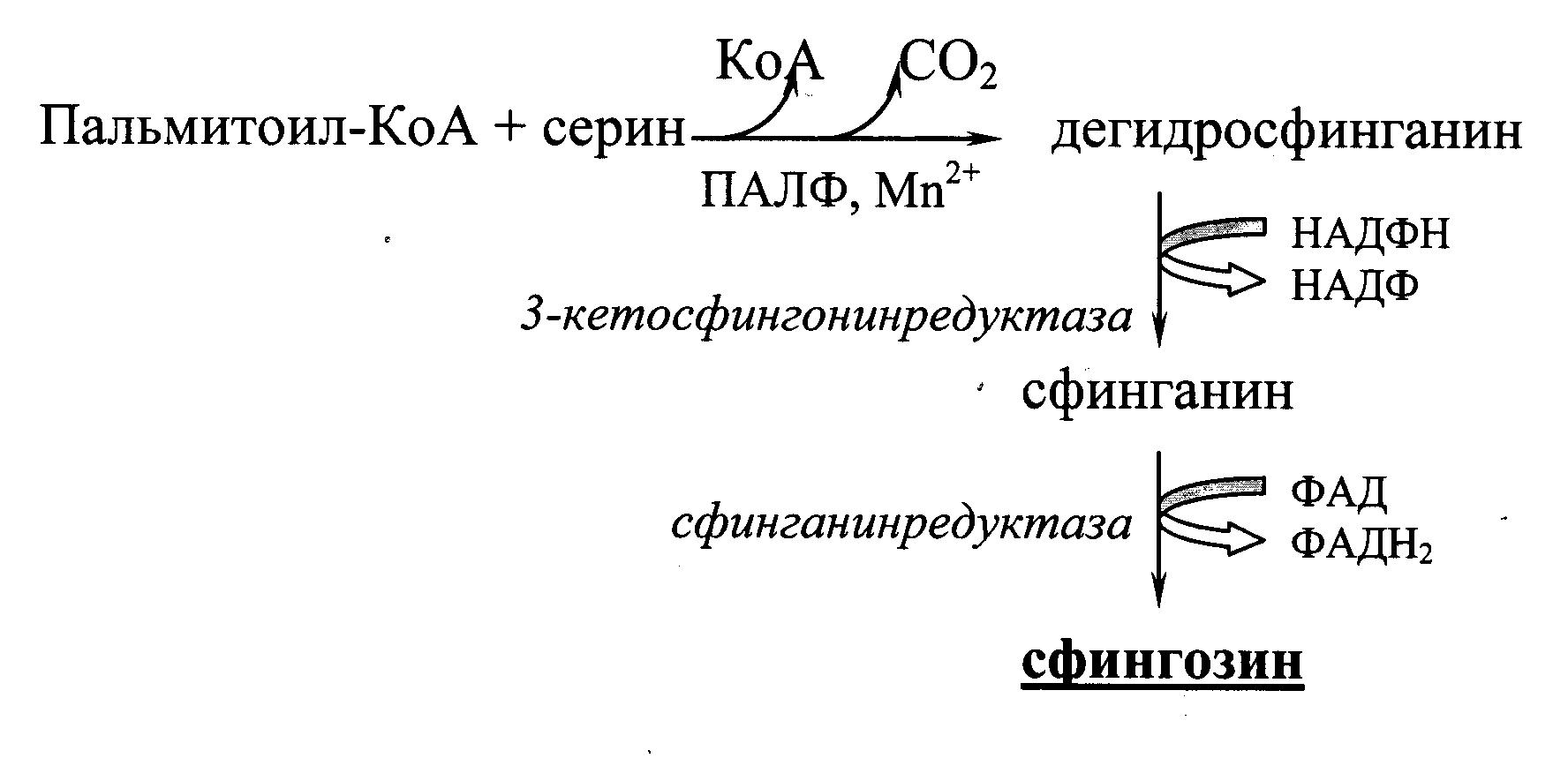
Рис. 21.1. Схема образования сфингозина
Сфингозин подвергается ацилированию (присоединение остатка жирной кислоты), в результате образуется церамид, из которого могут синтезироваться цереброзиды, ганглиозиды, сульфатиды и сфингомиелин (Рис.21.2.).
Катаболизм сфингомиелинов и гликолипидов происходит в лизосомах. Исключительно важный аспект этого процесса заключается в существовании более десяти специфических лизосомных болезней накопления – сфинголипидозов. Сфинголипидозы обычно являются причиной умственной отсталости и ведут к смерти в раннем возрасте, так как происходит поражение клеток нервной ткани, где сконцентрированы гликолипиды.
В распаде сфингомиелинов (Рис.21.2.) участвует сфингомиелиназа, отщепляющая фосфохолин. Генетический дефект сфингомиелиназы – причина болезни Нимана‑Пика. Дети с таким дефектом погибают в раннем возрасте. Симптомы болезни: накопление сфингомиелина в лизосомах, умственная отсталость, гепатоспленомегалия.
Сложные молекулы гликолипидов расщепляются в результате последовательных реакций гидролиза до глюкозы, галактозы, церамида и других метаболитов.
Генетические дефекты любого из ферментов, обеспечивающих катаболизм этого класса липидов, ведут к развитию заболеваний, среди которых можно назвать:
1. болезнь Гоше – следствие дефекта b‑глюкозидазы, при которой наблюдаются гепатоспленомегалия и умственная отсталость;
2. болезнь Тея‑Сакса – следствие дефекта b‑гексозаминидазы, для которой характерны умственная отсталость и слепота;
3. генерализованный ганглиозидоз, вызываемый снижением активности b‑галактозидазы, также ведущий к умственной отсталости.
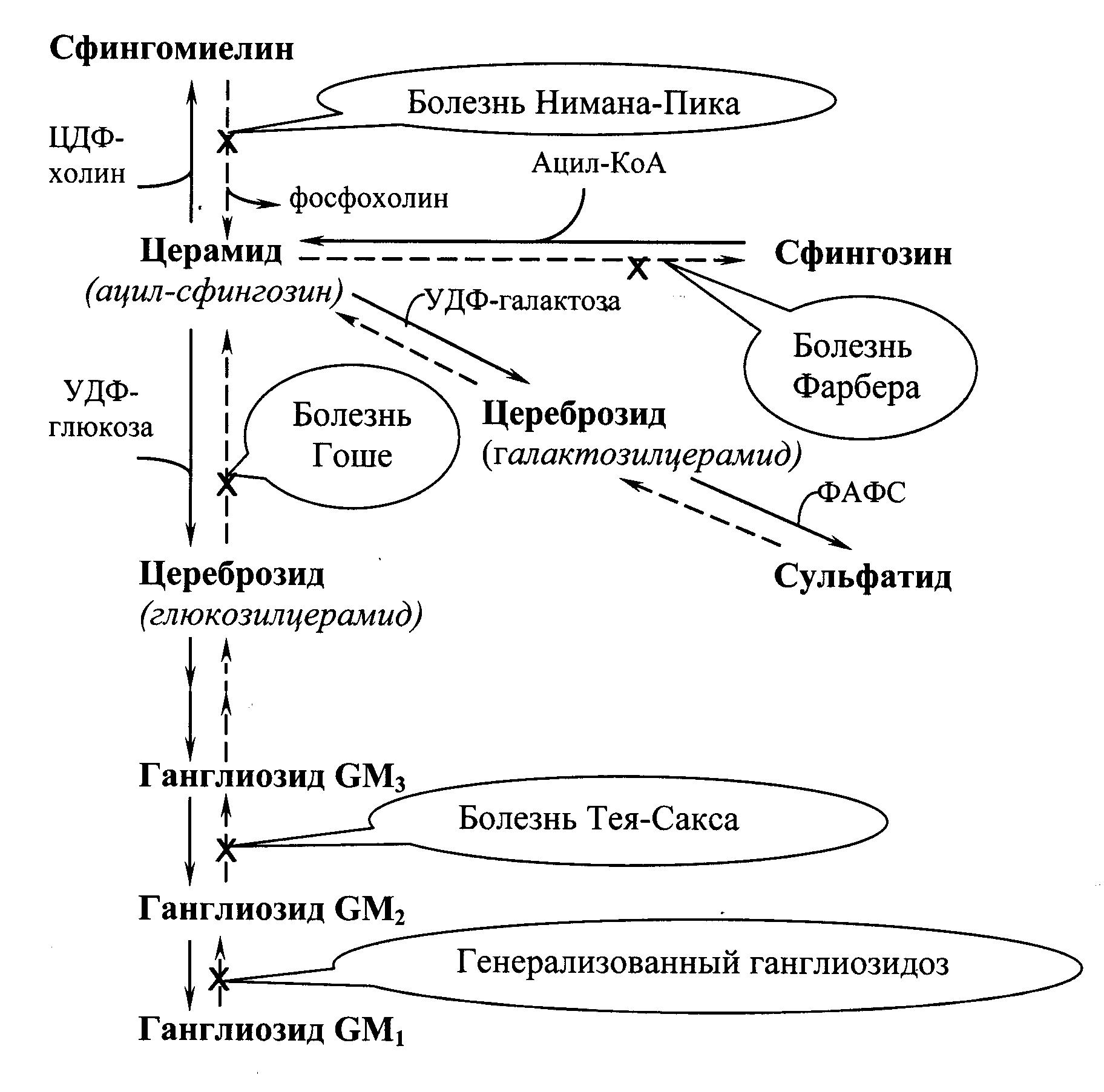
Рис.21.2. Биосинтез ( ) и распад ( ) сфинголипидов с указанием биохимических нарушений при сфинголипидозах.
Расщепление церамида до сфингозина и жирной кислоты осуществляется церамидазой. Генетический дефект этого фермента приводит к развитию болезни Фарбера с летальным исходом в раннем возрасте. При данной патологии в лизосомах накапливается церамид, наблюдается гепатоспленомегалия, умственная отсталость и поражения суставов.
Дата добавления: 2016-01-30; просмотров: 1306;