Классификация медицинской электронной аппаратуры 5 страница
Кривые течения – графики зависимости напряжения сдвига от скорости сдвига.
На рис. 5.2 кривая 1 соответству  ет ньютоновской жидкости. Видим, что ньютоновская жидкость начинает течь при самых небольших скоростях сдвига. Кривая 2 соответствует неньютоновской жидкости, т.е. вязко-пластичной жидкости. Здесь жидкость начинает течь только при
ет ньютоновской жидкости. Видим, что ньютоновская жидкость начинает течь при самых небольших скоростях сдвига. Кривая 2 соответствует неньютоновской жидкости, т.е. вязко-пластичной жидкости. Здесь жидкость начинает течь только при
| Рис. 5.2 | 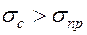 . . 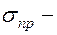 предел предел
|
текучести. Это связано с тем, что для приведения в движение агрегатов – монетных столбиков нужны дополнительные усилия в сравнении с ньютоновскими жидкостями.
Зависимость коэффициента вязкости крови от различных физических факторов.
1)  от градиента скорости (скорости сдвига)
от градиента скорости (скорости сдвига)
1 –эффективная вязкость,
2 – коэффициент вязкости ньюто-новской жидкости (взяли  равный кажущейся вязкости крови – предельной вязкости).
равный кажущейся вязкости крови – предельной вязкости).
Рис. 5.3
Из графика видно, что с увеличением скорости сдвига, т.е. с увеличением  , эффективная вязкость крови резко падает и при γ > 100 с-1 вязкость крови становится равной некоторому предельному значению, остающемуся далее неизменным как у ньютоновской жидкости. Это происходит оттого, что агрегаты эритроцитов с увеличением градиента скорости распадаются, следовательно, кровь находится под напряжением .
, эффективная вязкость крови резко падает и при γ > 100 с-1 вязкость крови становится равной некоторому предельному значению, остающемуся далее неизменным как у ньютоновской жидкости. Это происходит оттого, что агрегаты эритроцитов с увеличением градиента скорости распадаются, следовательно, кровь находится под напряжением .
Предельное значение вязкости крови называют кажущейся вязкостью.
2) от гематокритного показателя (гематокрита)  ,
,
т.е. от концентрации эритроцитов в крови:

С увеличением гематокритного показателя вязкость крови падает, т.к. увеличивается число комплексов и увеличиваются их раз
меры.
Рис. 5.4
3) от температуры
С увеличением температуры вязкость уменьшается – уменьша-
ются силы взаимодействия между молекулами эритроцитов, уменьшаются размеры агрегатов.

Рис. 5.5
В пределах температур от 100С до 380С вязкость крови имеет экспоненциальную зависимость от температуры


При температурах меньше 100С и выше 380С зависимость очень сложная, что связано с процессами, приводящими к изменению свойств крови.
3) от диаметра сосуда, по которому течет кровь
С увеличением диаметра сосуда вязкость крови увеличи-вается. В сосудах диаметром меньше 50 мкм этот эффект проявляется столь сильно, что может маскировать зависимость вязкости крови от скорости сдвига и от гематокритного показателя.

Рис. 5.6
Одно из объяснений этого эффекта такое. В сосудах с боль
шим  диаметром вся стопка движется целиком, увеличивая вязкость крови. В капиллярах эти агрегаты распадаются, эритроциты могут даже деформироваться - размеры частиц уменьшаются, вязкость падает.
диаметром вся стопка движется целиком, увеличивая вязкость крови. В капиллярах эти агрегаты распадаются, эритроциты могут даже деформироваться - размеры частиц уменьшаются, вязкость падает.
Физические модели кровообращения.
1) Гидродинамическая
| а) | б) | в) |

| 
| 
|
Рис. 5.7
Такую модель представляют в виде замкнутой, т.е. не имеющей сообщений с атмосферой системы трубок с эластич-ными стенками. Изначально считаем, что эта система заполнена жидкостью. Движение жидкости в ней происходит под действием ритмично работающего насоса в виде груши. Широкие трубки – аналог аорты и артерий, резкие разветвления – аналог периферической системы – каппиляров.
При сжатии груши, содержащийся в ней объём жидкости, проталкивается через отверстие в клапане 1 в систему трубок со стороны трубки А. Затем клапан 1 закрывается. В трубке уже была жидкость под некоторым давлением. Поэтому этот объём жидкости вызывает увеличение давления в широкой трубке, она расширяется. Затем под действием эластичных свойств трубка постепенно сжимается и жидкость проталкивается в следующее звено системы в сторону Б. Когда жидкость подойдет к трубке Б, то через отверстие клапане 2 она попадает снова в насос. Благодаря эластичности стенок трубок и ритмичной работы насоса течение жидкости в системе остается равномерным.
В этой системе имеется постепенное и множественное разветвление трубок, особенно в средней её части (множество параллельных трубок малого сечения). Общее число их такое большое, что скорость течения жидкости снижается здесь до нуля. Внутреннее трение в пристеночных слоях этих трубок очень велико, поэтому именно эта часть системы представляет наибольшее сопротивление течению жидкости и создает наибольшее падение давления.
2) Электрическая модель.
На основе рассмотренной гидродинамической механической модели можно построить электрическую модель кровообращения.Построена электрическая модель, исходя из следующих соображений. Жидкость течет, ток тоже течет. Перепад давлений вызывает ток жидкости, а разность потенциалов вызывает электрический ток.

На рисунке: 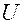 - источник переменного несинусоидального переменного напряжения, который служит аналогом сердца. Выпрямитель (диод)
- источник переменного несинусоидального переменного напряжения, который служит аналогом сердца. Выпрямитель (диод) 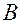 , пропускающий ток только в одном направлении, служит аналогом клапана. Конденсатор
, пропускающий ток только в одном направлении, служит аналогом клапана. Конденсатор 
| Рис. 5.8 | служит аналогом упругого резервуа |
ра (аорты и артерий). Действительно. Коденсатор служит для накопления заряда. В первые полпериода накапливает заряд, а во вторые полпериода разражается через сопротивление 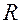 . Участки упругих резервуаров (аорта, артерии), обладающие элласичностью, можно рассматривать как емкость для крови. Вязкостное сопротивление, большее у переферичесих сосудов, можно представить в виде резистора .
. Участки упругих резервуаров (аорта, артерии), обладающие элласичностью, можно рассматривать как емкость для крови. Вязкостное сопротивление, большее у переферичесих сосудов, можно представить в виде резистора .
Электрическая модель широко применяется в теоретической медицине, т.к. распределение импульсов электрического потенциала и тока в электрических цепях хорошо изучены и легко описываются математически.
Закономерности выброса и распространения крови
в большом круге кровообращения.
Для рассмотрения этого вопроса вернемся к гидродинамической модели кровообращения.
Начальное давление, необходимое для продвижения крови по всей сосудистой системе, создается работой сердца.
При каждом сокращении левого желудочка сердца в аорту, уже заполненную кровью под соответствующим давлением, выталкиваться некоторый объем крови, называемый ударным объёмом (  65 – 70 мл). Затем сердечный клапан закрывается.
65 – 70 мл). Затем сердечный клапан закрывается.
Поступивший в аорту дополнительный объём крови повышает в ней давление и растягивает её стенки:

Таким образом, кровь по аорте распространяется как волна. Эта волна повышенного давления, называемого систолическим, вызывает колебания сосудистых стенок. Колебания далее распространяются вдоль крупных артерий в виде упругой волны. Эта волна повышенного давления, вызванная выбросом крови из левого желудочка во время систолы, называется пульсовой волной. Скорость пульсовой волны 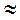 5 – 10 м/c.
5 – 10 м/c.
В период расслабления сердечной мышцы (диастолы) стенки аорты постепенно сокращаются до исходного состояния и проталкивают поступивший объём крови в более крупные артерии. В них, в свою очередь, стенки растягиваются и проталкивают кровь в последующие звенья системы.
Т.к. скорость крови 5 – 10 м/c, то за время систолы ( 0,3 с) она должна распространиться на расстояние 1,5 – 3 метра, что больше, чем расстояние от сердца до конечностей. Это означает, что пульсовая волна достигает конечностей раньше, чем начинается спад давления в аорте, т.е. прежде, чем начнется диастола. В результате распространения пульсовой волны кровь течет непрерывно.
Если графически представить пульсовую волну как гармоническую, то в аорте эта волна имеет график вид как на рис.
5.9 а, а в артериолах - как на рис. 5.9 б.
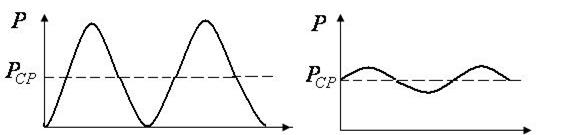
| Рис. 5.9 а | Рис. 5.9 б |
Амплитуда колебаний в аотре больше, чем в артериолах. Вязкость крови и упруго-вязкие свойства стенок сосудов уменьшают амплитуду колебаний пульсовой волны.
Течение крови в сосудистой системе в нормальных условиях носит ламинарный характер. Турбулентным оно может стать при резком сужении просвета.
Количество крови, протекающее через поперечное сечение участка сосудистой системы в 1 секунду, называется объёмной скоростью кровотока и может быть рассчитана по закону Гагена-Пуазейля
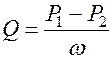 .
.
Но расчет этот приближенный, т.к.  зависит не только от
зависит не только от  и
и  , но и от потерь энергии при деформации эллчстичных стенок сосудов, от завихрений в местах разветвлений.
, но и от потерь энергии при деформации эллчстичных стенок сосудов, от завихрений в местах разветвлений.
Сопротивление току крови и падение давления на разных участках сосудистой системы различны (рис.5.10)

Рис. 5.10
Оно зависит от общего сечения (общего просвета) и от числа сосудов в разветвлении. Наибольшее падение происходит в артериолах (до 50 -ти % от начального). Это связано с тем, что число артериол в сотни раз больше, чем крупных артерий при сравнительно небольшом увеличении общего просвета сосудов. Поэтому потери давления от пристеночного трения достаточно большие. Общее число капилляров ещё больше, поэтому даже при небольшой их длине их падение давления в них значительно, но меньше, чем в артериолах.
В венах падение давления незначительно. Это связано с тем, что площадь сечения венозных сосудов в среднем в 2 раза больше, чем в артериях. Имеется участок, в котором давление оказывается ниже атмосферного. Этот участок соответствует движению крови под влиянием присасывающего действия грудной клетки при вдохе.
На рисунке представлен также график изменения скорости крови при прохождении её по разным сосудам. Как видим из рисунка в артериях скорость крови постоянна. В артериолах она падает и в капиллярах становится равной нулю. В венах скорость крови повышается.
Работа и мощность сердца.
Работа, совершаемая сердцем, складывается в основном из работы при сокращении желудочков сердца, главным образом ле-вого желудочка. Работа правого желудочка равна 0,15 –0,2 АЛ.Ж..
Работа сердечной мышцы при каждом сокращении левого желудочка затрачивается на сообщение объёму выталкиваемой крови энергии, необходимой для его продвижения по всему кругу кровеобращения. Эта энергия состоит из потенциальной энергии давления, которое должно быть создано вначале для преодоления по всему её пути, и кинетической энергии для сообщения массе крови необходимой скорости движения:


Здесь:  среднее давление, под которым кровь проталкивается в аорту (
среднее давление, под которым кровь проталкивается в аорту (  );
); 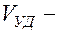 ударный объём крови (
ударный объём крови (  70 мл);
70 мл); 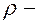 плотность крови (1,05·103 кг/м3);
плотность крови (1,05·103 кг/м3);  скорость крови в аорте ( 0,5 м/с в состоянии покоя)
скорость крови в аорте ( 0,5 м/с в состоянии покоя)
При тех условиях, что я здесь привела, получаем за одно сокращение сердца работа левого желудочка:

а работа сердца
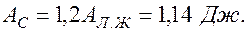
Время сокращения желудочков  Тогда мощность сердца, развиваемая за одно сокращение
Тогда мощность сердца, развиваемая за одно сокращение
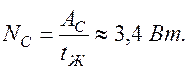
За одну минуту число сокращений сердца 60. Поэтому
| за 1 минуту | 
|
| за сутки | 
|
Основываясь на этих данных, был создан аппарат искусственного кровеобращения. Он представляет собой насосную систему (искусственное сердце) и оксигенератор (искусственные легкие) для обеспечения насыщения крови кислородом.
Измерение давления крови.
Лабораторный метод измерения давления крови мы уже рассмотрели при изучении вопроса “Следствия из уравнения Бернулли”
Бескровный метод измерения артериального давления – метод Короткова, основанный на выслушивании звуков, возникащих при прохождении крови через сжатую манжетой артерию.
На предплечье исследуемог укрепляют манжету и прощупывают пульс плечевой артерии несколько выше локтевого сгиба, прикладывают к этому месту фонендоскоп.. Нагнетают в манжету воздух до давления на 10  20 мм.рт.ст.выше, чем то, при котором перестает прощупываеться пульс. Затем, медленно вращая выпускной винт нагнетателя, постепенно снижают давление в манжете, прислушиваясь к звукам, появляющимся в фонендоскопе.
20 мм.рт.ст.выше, чем то, при котором перестает прощупываеться пульс. Затем, медленно вращая выпускной винт нагнетателя, постепенно снижают давление в манжете, прислушиваясь к звукам, появляющимся в фонендоскопе.  Соотношение между изменением давления
Соотношение между изменением давления  в мажете и “тонами Короткова” показано на рис. 5.12.
в мажете и “тонами Короткова” показано на рис. 5.12.  систолическое давление (в норме 100 120 мм.рт.ст.);
систолическое давление (в норме 100 120 мм.рт.ст.);
 диастолическое давление (в норме 70 80 мм.рт.ст.)
диастолическое давление (в норме 70 80 мм.рт.ст.)
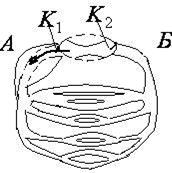
Пока артерия полностью сжата, никаких звуков не прослушивается. При снижении в манжете давления начинают прослушиваться отчетливые тоны, называемые начальными (а на рис.12). Появляется пульс на лучевой артерии. Эти тоны обусловлены вибрацией стенок артерии непосредственно за манжетой под действием толчков крови, которые прорываются через сжатый манжетой участок сосуда только в моменты систолысердца (максимальное давление).
Показания манометра при первом появлении тонов соответствуют систолическому давлению. При дальнейшем понижении давления в манжете появляются шумы (б на рис.12) интенсивности больше, чем у тонов. Эти шумы обусловлены турбулентным движением крови через частично сдавленный манжетой участок артерии. Затем шумы стихают и вновь прослушиваются тоны (в на рис.12). Эти тоны быстро стихают и звуковые явления исчезают. Это происходит при полном восстановлении просвета артерии и установлении нормального ламинарного течения крови. Показания манометра в момент резкого ослабления последовательных тонов соответствуют минимальному или даистолическому давлению.
В последнее время появились электронные манометры. В них используются параметрические датчики, в которых давление крови меняет какие-либо параметры датчика. В них давление высвечивается прямо на экранчике.
Лекция 6.
БИОЛОГИЧЕСКИЕ МЕМБРАНЫ
Значение и функции биологических мембран. Строение мембран.
Структурной и функциональной единицей живого организма является клетка, которой присущи все основные жизненные функции. Все клетки состоят из цитоплазмы, окруженной плазматической мембраной. В цитоплазме находятся ядро, органоиды клетки (метахондрии, липосомы, рибосомы и др.).
Мембраны окружают всю цитоплазму и отграничивают её от окружающей среды. Проникновение веществ в клетку и из клетки в большей степени зависит от свойств мембран. Мембраны находятся и внутри клетки, образуя оболочки всех органоидов и включений клетки.
Толщина мембраны порядка нескольких нанометров, (≈ 10-9 м.), поэтому в оптический микроскоп её разглядеть нельзя, но зато она хорошо рассматривается в электронном микроскопе.
В 1935 году Даниэлли и Давсон предложили модель строения мембраны, которая не претерпела существенных изменений до нашего времени. Согласно этой модели основу мембраны составляют два слоя фосфолипидов (рис. 6.1).
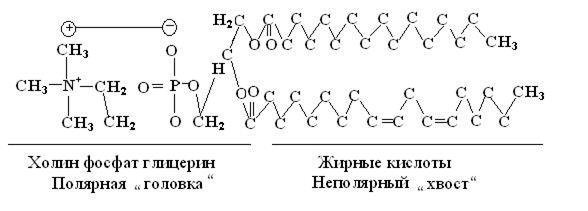
Рис.6.1
Молекула фосфолипида имеет полярную “головку” и неполярный “ хвост”. Два слоя молекул фосфолипидов расположены перпендикулярно поверхности мембраны (рис.6.2). Гидрофильные концы молекул (полярные головки) способны

Рис.6.2
взаимодействовать с дипольными молекулами воды и формировать гидратные оболочки. Поэтому гидрофильные (полярные) концы молекул липидов направлены наружу мембраны. Гидрофобные (неполярные) концы молекул направлены вглубь мембраны - они не могут присоединять молекулы воды. Молекулы фосфолипидов адсорбируют молекулы белков.
Моделей мембран высказывалось очень много, но ни одна из них не стала общепринятой.
В настоящее время наибольшее распространение получила жидкомозаичная модель, предложенная в 1972 году Синджером и Никольсоном. В основе этой модели лежит также двухслойная липидная мембрана. Эта фосфолипидная основа представляет собой как бы двумерный растворитель, в котором плавают более или менее погруженные белки

Рис. 6.3
За счет указанных белков полностью или частично осуществляются специфические функции мембран: проницаемость, активный транспорт через мембрану, генерация электрической разности потенциалов и др.
Мембраны не являются неподвижными структурами. Липиды и белки обмениваются местами и перемещаются вдоль плоскости (латеральная диффузия) и поперек плоскости мембран (диффузия флип-флоп) .Латеральная диффузия сопровождается высокой подвижностью липидов, а диффузия флип-флоп – низкой подвижностью, т.е. обмен местами липидов, находящихся на разных сторонах мембраны, является маловероятным процессом.
Физиологические функции мембран.
1) Делят клетку на отдельные участки, фазы, в которых протекают различные биохимические реакции, которых в клетках протекает до тысячи видов.
2) Принимают участие во всех процессах обмена веществ, которые обусловливают жизнедеятельность клетки. Это происходит потому, что мембранные структуры в организмах составляют огромную поверхность (до десятков тысяч квадратных метров). Поэтому биохимические реакции протекают в основном на поверхности мембран, обеспечивая обмен веществ.
3) Координируют и регулируют биохимические и биофизические процессы в клетках. Мембраны являются своеобразным устройством, воспринимающим сигналы извне и преобразующим их в команды, регулирующие обмен веществ в клетке.
4) Клеточные мембраны обеспечивают адгезию (сцепление) клеток друг с другом, что обеспечивает существование ткани. Адгезия осуществляется специализированными структурами на клеточных мембранах (иногда это просто как механическое сцепление типа”гнездо-шип", иногда химическая связь, иногда- склеивающее цементнообразное вещество”).
2.Физические свойства мембран.
1. Измерение подвижности молекул мембраны и диффузия частиц через мембрану свидетельствуют о том, билипидный слой ведет себя подобно жидкости. Но, с другой стороны, мембрана есть упорядоченная структура, свойственная кристаллам. Эти два факта заставляют думать, что фосфолипиды в мембране находятся в жидкокристаллическом состоянии.
2. Вязкость липидного слоя мембраны на 2 порядка (в 100 раз) выше, чем у воды. От вязкости липидного слоя зависит скорость поступательного и вращательного движения молекул (в том числе мембранных белков и ионов), т.е. вязкость определяет подвижность ионов и молекул, проникающих через мембрану (проницаемость мембраны), а, следовательно, регулирует скорость протекающих в мембранах ферментативных реакций.
3. Поверхностное натяжение липидного слоя на 2-3 порядка (100-1000 раз) меньше, чем у воды. Значит поверхностная энеогия липидного слоя незначительна 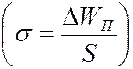 , что облегчает диффузию веществ через мембрану.
, что облегчает диффузию веществ через мембрану.
4. При изменении температуры в мембране наблюдаются фазовые переходы: плавление липидов при нагревании и кристаллизация при охлаждении. Структура молекул в жидком и кристаллическом состояниях различна. В жидкой фазе молекулы фосфолипидов могут образовывать полости (кинки), в которые способны внедряться молекулы диффундирующего вещества.
5. Двойной фосфолипидный слой уподобляепт мембрану конденсатору. Электроёмкость 1 мм2 мембраны составляет 5-13 нФ.
Модели искусственных мембран для изучения свойств мембран.
1. Частокол Ленгмюра – монослой фосфолипидов. Модель создана в 1917 году.
На поверхность воды наносят каплю растворенных  в каком-либо растворителе фосфолипидов или жирных кислот. После распределения их молекул на поверхности воды и испарения расворителя на поверхности воды остается пленка. Адсорбированные молекулы липидов располагаются перпендикулярно поверхности воды.
в каком-либо растворителе фосфолипидов или жирных кислот. После распределения их молекул на поверхности воды и испарения расворителя на поверхности воды остается пленка. Адсорбированные молекулы липидов располагаются перпендикулярно поверхности воды.
2. Липосомы – широко распространенная модель. Это мель-чайшие пузырьки, состоящие из билипидной мембраны. Получаются при обработке смеси воды и фосфолипидов ультразвуком. На ней хорошо изучать влияние состава фосфолипидов на свойства мембран или влияние мембран на свойства втсраеваемых белков.

3. Билипидная мембрана. Берется два водных раствора, разде-
 ленных тефлоновой перегородкой
ленных тефлоновой перегородкой
с отверствием. Отверствие заполняют фосфолипидом, растворенным в гексане. Когда растворитель и изли-шек липида растекаются по тефлону, в отверстии образуется бислой, толщиной в несколько нанометров.
На такой мембране удобно изучать проницаемость мембраны для разных агентов, измерять сопротивление мембраны или генерируемый на ней потенциал.
Методы исследования мембран
1. Микрокалориметрия. Изучаются фазовые переходы на каком-либо участке мембраны и оценивают размеры “кооперативных единиц”, т.е. число молекул, участвующем в фазовом переходе. Для этого измеряют теплоемкость 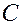 суспензии фосфолипидов при разных температурах в области фазового перехода. Например, для синтетического липида дистеароил-фосфатидил-холина графики зависимости от
суспензии фосфолипидов при разных температурах в области фазового перехода. Например, для синтетического липида дистеароил-фосфатидил-холина графики зависимости от 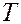 , где - абсолютная температура выглядят так
, где - абсолютная температура выглядят так


В области фазового перехода при 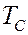 происходит резкое возрастание теплоемкости. Площадь под кривыми соответствует количеству теплоты
происходит резкое возрастание теплоемкости. Площадь под кривыми соответствует количеству теплоты 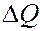 , поглощаемого при переходе из твердого состояния в жидкое. Зная массу
, поглощаемого при переходе из твердого состояния в жидкое. Зная массу 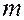 фосфолипида в пробе, можно рассчитать энтальпию плавления
фосфолипида в пробе, можно рассчитать энтальпию плавления

и энтропию при плавлении
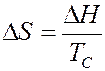 .
.
Энтрапия 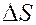 возрастает с ростом длины углеводородной цепи жирных кислот, причем на каждую метиленовую группу приходится примерно постоянное увеличение энтропии при фазовом переходе. Т.о. мы можем судить о длине куглеводородной цепи липидов.
возрастает с ростом длины углеводородной цепи жирных кислот, причем на каждую метиленовую группу приходится примерно постоянное увеличение энтропии при фазовом переходе. Т.о. мы можем судить о длине куглеводородной цепи липидов.
Площадь под кривой от  до
до 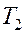

пропорциональна количеству молекул уже претерпевших фазовый переход.
2. ИК - спектроскопия.
Этот метод основан на замечательном свойстве природы: молекулы каждого вещества имеют индивидуальные, специфические спектры поглощения. Молекулярные спектры позволяют
исследовать состав мембран, строение молекул в мембране и характер межмолекулярных взаимодействий.
3. Люминесцентный анализ.
Производят на спектрофлюоиметрах.
Молекулы белков обладают флюоресценцией. Параметры флюоресценции чувствительны к структуре окружения флюоресцирующей молекулы. Поэтому по люминесценции можно изучать химические превращения и межмолекулярные взаимодействия.
Для исследования мембран к мембранным системам добавляют флюоресцирующие молекулы: флюоресцентные зонды, если молекула образует нековалентную связь с мембраной , или флюоресцентные метки, если молекула образует химическую связь с мембраной.
При освещении мембранной системы с флюоресцирующей молекулой поляризованным светом люминесценция оказывается также поляризованной. Если флюоресцентный зонд неподвижен (например, раствор заморожен), то степень поляризации 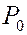 будет мексимальной. Однако, если зонд или метка находятся в жидком растворе, то он за время возбужденного состояния успевает переориентироваться (перестроится), а поэтому изменится и степень поляризации люминесценции до величины
будет мексимальной. Однако, если зонд или метка находятся в жидком растворе, то он за время возбужденного состояния успевает переориентироваться (перестроится), а поэтому изменится и степень поляризации люминесценции до величины  . Изменение степени поляризации зависит от вязкости окружающих зонд молекул. Т.о люминесцентный анализ позволяет определить вязкость мембран, а также конформационные перестройки в белках и мембранах.
. Изменение степени поляризации зависит от вязкости окружающих зонд молекул. Т.о люминесцентный анализ позволяет определить вязкость мембран, а также конформационные перестройки в белках и мембранах.
Дата добавления: 2016-01-20; просмотров: 1189;