Поток энергии в экосистемах
Поддержание жизнедеятельности организмов и круговорот вещества в экосистемах возможны только за счет постоянного притока энергии (рис. 146). В конечном счете вся жизнь на Земле существует за счет энергии солнечного излучения, которая переводится фотосинтезирующими организмами в химические связи органических соединений. 4етеротрофы получают энергию с пищей. Все живые существа являются объектами питания других, т. е. связаны между собой энергетическими отношениями. Пищевые связи в сообществах – это механизмы передачи энергии от одного организма к другому. В каждом сообществе трофические связи переплетены в сложную сеть. Организмы любого вида являются потенциальной пищей многих других видов. Врагами тлей, например, служат личинки и жуки божьих коровок, личинки мух-сирфид, пауки, насекомоядные птицы и многие другие. За счет дубов в широколиственных лесах могут жить несколько сотен форм различных членистоногих, фитонематод, паразитических грибков и т. п. Хищники обычно легко переключаются с одного вида жертв на другой, а многие, кроме животной пищи, способны потреблять в некотором количестве и растительную. Таким образом, трофические сети в биоценозах очень сложные и создается впечатление, что энергия, поступившая в них, может долго мигрировать от одного организма к другому.
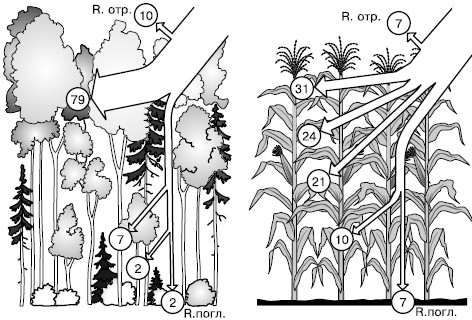
Рис. 146. Распределение радиации (в кружочках – %) в бореальном смешанном лесу и в посевах кукурузы (по В. Лархеру, 1978): R. отр. – радиация, отраженная от поверхности насаждения; R. погл. – радиация, поглощенная почвой
На самом деле путь каждой конкретной порции энергии, накопленной зелеными растениями, короток. Она может передаваться не более чем через 4–6 звеньев ряда, состоящего из последовательно питающихся друг другом организмов. Такие ряды, в которых можно проследить пути расходования изначальной дозы энергии, называют цепями питания (рис. 147).
Место каждого звена в цепи питания называют трофическим уровнем. Первый трофический уровень – это всегда продуценты, создатели органической массы; растительноядные консументы относятся ко второму трофическому уровню; плотоядные, живущие за счет растительноядных форм, – к третьему; потребляющие других плотоядных – соответственно к четвертому и т. д. Таким образом, различают консументов первого, второго и третьего порядков, занимающих разные уровни в цепях питания. Естественно, что основную роль при этом играет пищевая специализация консументов. Виды с широким спектром питания могут включаться в пищевые цепи на разных трофических уровнях. Так, например, человек, в рацион которого входит как растительная пища, так и мясо травоядных и плотоядных животных, выступает в разных пищевых цепях в качестве консумента первого, второго и третьего порядков. Виды, специализированные на растительной пище, например тли, зайцеобразные, копытные, всегда являются вторым звеном в цепях питания.
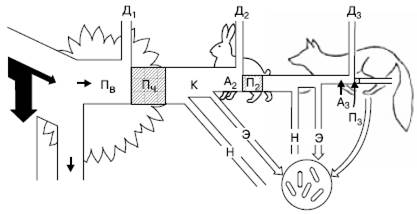
Рис. 147. Поток энергии через три уровня простой пищевой цепи (по П. Дювиньо и М. Тангу, 1968): Пв – продукция валовая; Пч – продукция чистая; К – продукция, использованная на корм; А2, А3 – корм, ассимилированный консументами; н – неиспользованная часть продукции; П2 – вторичная продукция (травоядные); П3 – прирост хищников; Д1-Д3 – траты энергии на обмен веществ (траты на дыхание) на разных уровнях пищевой цепи
Энергетический баланс консументов складывается следующим образом. Поглощенная пища обычно усваивается не полностью. Неусвоенная часть вновь возвращается во внешнюю среду (в виде экскрементов) и в дальнейшем может быть вовлечена в другие цепи питания. Процент усвояемости зависит от состава пищи и набора пищеварительных ферментов организма. У животных усвояемость пищевых материалов варьирует от 12–20 % (некоторые сапрофаги) до 75 % и более (плотоядные виды). Ассимилированная организмом пища вместе с запасом в ней энергии расходуется двояким образом. Большая часть энергии используется на поддержание рабочих процессов в клетках, а продукты расщепления подлежат удалению из организма в составе экскретов (мочи, пота, выделений различных желез) и углекислого газа, образующегося при дыхании. Энергетические затраты на поддержание всех метаболических процессов условно называют тратой на дыхание, так как общие их масштабы можно оценить, учитывая выделение СО2 организмом. Меньшая часть усвоенной пищи трансформируется в ткани самого организма, т. е. идет на рост или откладывание запасных питательных веществ, увеличение массы тела. Эти отношения сокращенно можно выразить формулой:
Р = П + Д + Н,
где Р – рацион консумента, т. е. количество пищи, съедаемой им за определенный период времени; П – продукция, т. е. траты на рост; Д – траты на дыхание, т. е. поддержание обмена веществ за тот же период; Н – энергия неусвоенной пищи, выделенной в виде экскрементов.
Передача энергии в химических реакциях в организме происходит, согласно второму закону термодинамики, с потерей части ее в виде тепла. Особенно велики эти потери при работе мышечных клеток животных, КПД которых очень низок. В конечном счете вся энергия, использованная на метаболизм, переходит в тепловую и рассеивается в окружающем пространстве.
Траты на дыхание во много раз больше энергетических затрат на увеличение массы самого организма. Конкретные соотношения зависят от стадии развития и физиологического состояния особей. У молодых траты на рост могут достигать значительных величин, тогда как взрослые используют энергию пищи почти исключительно на поддержание обмена веществ и созревание половых продуктов. Интенсивность питания снижается с возрастом. Так, ежесуточный рацион карпов массой от 5 до 15 г составляет почти 1/4 от массы их тела, у более крупных особей – от 150 до 450 г – всего 1/10, а у рыб массой 500–800 г – 1/16.
Коэффициент использования потребленной пищи на рост (К) рассчитывают как отношение продукции к рациону:
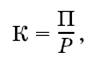
где П – траты на рост, Р – количество пищи, съеденной за тот же период.
Двупарноногие многоножки кивсяки в период роста, который продолжается до трех лет, тратят на рост от 6 до 25 % съеденной пищи при усвояемости в среднем 30 %. В последующем их масса стабилизируется. Кивсяки живут до 12 лет. В умеренном поясе они активны 4–5 месяцев в году. Особь, масса которой во взрослом состоянии 0,5 г, за свою жизнь потребляет 250–300 г опада (80–90 г абсолютно сухой массы). Так как кивсяки многократно линяют, часть усвоенной энергии идет на восстановление покровов. Таким образом, отношение съеденного в течение жизни корма к массе взрослого животного составляет 500–600: 1.
У такого гетеротермного животного, как малый суслик, который активен всего 2–2,5 месяца в году, это соотношение всего около 150: 1. Средний рацион суслика 30 г сухой массы растений (или в среднем 100 г сырой) при массе зверька 200 г и продолжительности жизни 4 года. Постоянно активным в течение года рыжим полевкам нужно гораздо больше энергии для поддержания жизнедеятельности. Взрослые зверьки массой 20 г съедают в среднем до 4 г сухого корма в день. При продолжительности жизни примерно в 24 месяца затрата кормов на жизнь одной особи составляет примерно 30 кг в сырой массе, что приблизительно в 1500 раз больше массы взрослого животного.
Таким образом, основная часть потребляемой с пищей энергии идет у животных на поддержание их жизнедеятельности и лишь сравнительно небольшая – на построение тела, рост и размножение. Иными словами, большая часть энергии при переходе из одного звена пищевой цепи в другое теряется, так как к следующему потребителю может поступить лишь та энергия, которая заключается в массе поедаемого организма. По грубым подсчетам, эти потери составляют около 90 % при каждом акте передачи энергии через трофическую цепь. Следовательно, если калорийность растительного организма 1000 Дж, при полном поедании его травоядным животным в теле последнего останется из этой порции всего 100, в теле хищника – лишь 10 Дж, а если этот хищник будет съеден другим, то на его долю придется только 1 Дж, т. е. 0,1 %.
Таким образом, запас энергии, накопленный зелеными растениями, в цепях питания стремительно иссякает. Поэтому пищевая цепь включает обычно всего 4–5 звеньев. Потерянная в цепях питания энергия может быть восполнена только поступлением новых ее порций. Поэтому в экосистемах не может быть круговорота энергии, аналогичного круговороту веществ (рис. 148). Экосистема функционирует только за счет направленного потока энергии, постоянного поступления ее извне в виде солнечного излучения или готовых запасов органического вещества.
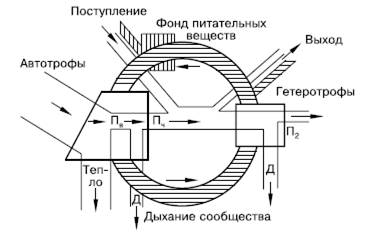
Рис. 148. Схема биогеохимического круговорота на фоне потока энергии (по Ю. Одуму, 1975): Пв – валовая продукция; Пч – чистая продукция; П2 – вторичная продукция; Д – траты энергии на обмен веществ (траты на дыхание); заштрихованная часть рисунка – круговорот вещества
Трофические цепи, которые начинаются с фотосинтезирующих организмов, называют цепями выедания (или пастбищными, или цепями потребления), а цепи, которые начинаются с отмерших остатков растений, трупов и экскрементов животных, – детритными цепями разложения. Таким образом, поток энергии, входящий в экосистему, разбивается далее как бы на два основные русла, поступая к консументам через живые ткани растений или запасы мертвого органического вещества, источником которого также является фотосинтез.
В разных типах экосистем мощность потоков энергии через цепи выедания и разложения различна: в водных сообществах большая часть энергии, фиксированной одноклеточными водорослями, поступает к питающимся фитопланктоном животным и далее – к хищникам и значительно меньшая включается в цепи разложения. В большинстве экосистем суши противоположное соотношение: в лесах, например, более 90 % ежегодного прироста растительной массы поступает через опад в детритные цепи (рис. 149).
А

Б
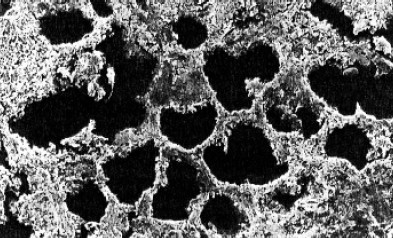
В
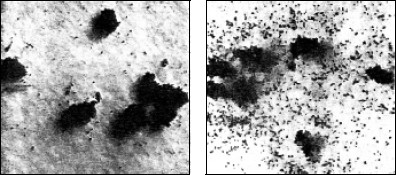
Рис. 149. Разложение листьев в лесной подстилке (деструкционный блок экосистемы):
А – поражение грибным мицелием;
Б – скелетирование животными-сапрофагами;
В – экскременты сапрофагов, входящие в состав гумусного слоя
Дата добавления: 2015-11-26; просмотров: 1004;