Бинокулярное зрение.
При взгляде на какой-либо предмет у человека с нормальным зрением не возникает ощущения двух предметов, хотя и имеется два изображения на двух сетчатках. Изображения всех предметов попадают на так называемые корреспондирующие, или соответственные, участки двух сетчаток, и в восприятии человека эти два изображения сливаются в одно. Надавите слегка на один глаз сбоку: немедленно начнет двоиться в глазах, потому что нарушилось соответствие сетчаток. Если же смотреть на близкий предмет, конвергируя глаза, то изображение какой-либо более отдаленной точки попадает на неидентичные (диспаратные) точки двух сетчаток. Диспарация играет большую роль в оценке расстояния и, следовательно, в видении глубины рельефа. Человек способен заметить изменение глубины, создающее сдвиг изображения на сетчатках на несколько угловых секунд. Бинокулярное слитие или объединение сигналов от двух сетчаток в единый нервный образ происходит в первичной зрительной коре. При бинокулярном зрении возникают содружественные движения глаз, т.е. в системе внешних координат глаза движутся в одном направлении – вверх, низ, влево, вправо. При перемещении, рассматриваемых предметов издали все ближе и ближе происходит конвергентное (конвергенция – схождение) движение глаз. Если близко расположенный предмет начинает удаляться все дальше и дальше, то зрительные оси расходятся (дивергенция глаз). Когда мы осматриваемся вокруг, то наши глаза совершают быстрые саккадические (скачкообразные) движения. Эти движения могут иметь микросаккадический характер, когда амплитуда движений составляет всего несколько угловых минут. При перемещении в поле зрения объектов возникают так называемые плавные следящие движения. Движения глаз могут быть в виде нистагма, когда чередуются медленные следящие движения и саккады. Подобные движения глаз могут быть, например, тогда, когда едущий в поезде человек фиксирует предметы, находящиеся за окном. В этом случае оба глаза совершают содружественные движения в направлении противоположном движению поезда и стремятся сохранить в поле зрения зафиксированный ранее объект. Когда этот объект исчезает с поля зрения саккадическое (быстрое) движение в обратном направлении (по ходу движения поезда) переводит взгляд на новую точку фиксации. Подобные движения глаз возникают также при вращении, но нистагм головы и глаз может продолжаться и после прекращения вращения. При чтении возникают саккады справа налево, а если текст вертикальный (китайский или японский), то и саккады имеют вертикальную форму. При чтении трудного для понимания текста или у детей возникают так называемые возвратные саккады, т.е. противоположные обычным направлениям движения глаз.
Величина предмета оценивается как функция величины изображения на сетчатке и расстояния предмета от глаза. В случае, когда расстояние до незнакомого предмета оценить трудно, возможны грубые ошибки в определении его величины.
ФИЗИОЛОГИЯ СЛУХА
Слух представляет собой такую функцию организма, которая неразрывно связана по своему происхождению со звуком. Как физическое явление звук представляет собой упругие волны, распространяющиеся в средах, существующих на земле, (в газах, жидкостях и твердых телах). Звуковые волны могут быть охарактеризованы целым рядом параметров: длиной волны, скоростью распространения в упругой среде, частотой колебаний, давлением, оказываемым на преграду, переносимой мощностью, явлениями отражения и т. д.. Звуковые волны не являются носителями больших мощностей, но нередко они возникают вместе с носителями больших мощностей, обладающих громадной знергией (взрыв, молния – гром и т. д.). Задача органа слуха состоит не просто в приеме звуковых колебаний, а в установлении местонахождения источника звука, его свойств. На основе начального элементарного анализа, а затем детального высшего анализа формируется адекватная ситуации поведенческая реакция. Звуковая волна распространяется в воздухе со скоростью 335 м/сек. Уровень звукового давления измеряют в децибелах (дБ), кроме того, в акустике учитывают частоту (Гц). Звуки, образованные одной частотой, называют тонами. Чистые тоны в повседневной жизни практически не встречаются, большинство звуков образовано наложением нескольких частот. Звук, состоящий из множества несвязанных между собой частот, называется шумом. Слуховая система является одним из важнейших' дистантных сенсорных систем человека и животных. Слуховой анализатор, так же как и другие анализаторы состоит из 3 основных отделов: воспринимающая (периферическая) часть, проводниковая часть (включает в себя слуховые нервы и подкорковые структуры) и корковый конец анализатора. Периферическая (воспринимающая) часть органа слуха делится на звукопроводящую (наружное и среднее ухо) и звуковоспринимающую (внутреннее ухо) части. Наружный слуховой проход проводит звуковые колебания к барабанной перепонке, которая отделяет наружное ухо от барабанной полости, или среднего уха. Барабанная перепонка представляет тонкую (0,1 мм) перегородку, в виде направленной внутрь воронки. Перепонка колеблется при действии звуковых колебаний. В заполненном воздухом среднем ухе находятся три косточки: молоточек, наковальня и стремечко, они передают колебания барабанной перепонки во внутреннее ухо. Рукоятка молоточка прочно соединена с барабанной перепонкой, другая сторона молоточка соединена с наковальней, передающей колебания стремечку. Благодаря особенностям геометрии слуховых косточек колебания барабанной перепонки передаются стремечку уменьшенной амплитуды, но увеличенной силы. Поверхность стремечка в 22 раза меньше барабанной перепонки, в результате во столько же раз усиливается его давление на мембрану овального окна. Поэтому действующие на барабанную перепонку, даже слабые звуковые волны, преодолевают сопротивление мембраны овального окна преддверия и приводят к колебаниям жидкости в улитке. Колебания барабанной перепонки облегчает также слуховая (евстахиева) труба, соединяющая среднее ухо с носоглоткой, давление в среднем ухе выравнивается с атмосферным. Кроме овального, есть еще круглое окно улитки, тоже закрытое мембраной. Колебания жидкости улитки, возникшие у овального окна достигают, не затухая, круглого окна улитки. В среднем ухе расположены две мышцы: напрягающие барабанную перепонку (m. tensor tympani) и стременная (m. stapedius). При сокращении первой усиливается натяжение барабанной перепонки и тем самым ограничивается амплитуда ее колебаний при сильных звуках, а вторая фиксирует стремечко и тем самым ограничивает его движения. Рефлекторное сокращение этих мышц наступает через 10 мс после начала сильного звука и зависит от его амплитуды. Этим внутреннее ухо автоматически предохраняется от перегрузок. При мгновенных сильных раздражениях (удары, взрывы и т. д.) этот защитный механизм может не успеть сработать, в результате чего возможны нарушения слуха (например, у взрывников и артиллеристов).
Во внутреннем ухе находится улитка, которая имеет слуховые рецепторы. Улитка это костный спиральный канал, имеющий 2,5 витка. Диаметр костного канала у основания улитки 0,04 мм, а на вершине ее — 0,5 мм. Костный канал улитки разделен двумя перепонками: более тонкой — преддверной (вестибулярной) мембраной (мембрана Рейсснера) и более плотной и упругой — основной мембраной. На вершине улитки обе эти мембраны соединяются, и в них имеется овальное отверстие улитки — helicotrema. Вестибулярная и основная мембраны разделяют костный канал улитки на три хода: верхний, средний и нижний.
Верхний канал улитки, или лестница преддверия (scala vestibuli - 1), у овального окна преддверия через овальное отверстие улитки (helicotrema) сообщается с нижним каналом улитки — барабанной лестницей (scala tympani - 18). Верхний и нижний каналы улитки заполнены перилимфой, напоминающей по составу цереброспинальную жидкость.
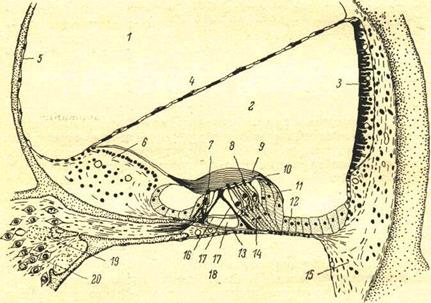
Рис. 51. Поперечный разрез улитки (Davis, 1961).
1 — scala veetlbuli (перилимфа); 2— scala media (эндолимфа); 3 — stria vascularis; 4 — рейснерова мембрана; 5— костная стенка улиткового канала; 6— Ilmbus; 7 — внутренние волосковые клетки; 8 — наружные волосковые клетки; 9 — текториальная мембрана; 10 — ретикулярная пластинка; 11 — клетки Гензена; I2 — клетки Клаудиуса; 13 — базилярная мембрана; 14 — клетки Дейтерса; I5— спиральная связка;16 — нервные волокна; 17— столбы и туннель Корти; 18 — scala tympani (перилимфа); 19 — интра-ганглионарный спиральный пучок, состоящий из эфферентных волокон и идущий во внутреннем костном канале (modiolus), составляющем ось улитки; 20 — клетки спирального ганглия.
Между верхним и нижним каналами проходит средний — перепончатый канал (scala media - 2). Полость этого канала не сообщается с полостью других каналов и заполнена эндолимфой. Эндолимфа содержит в 100 раз больше калия и в 10 раз меньше натрия, чем перилимфа, поэтому эндолимфа заряжена положительно по отношению к перилимфе.
Внутри среднего канала улитки на основной (базилярной) мембране расположен кортиев орган — рецепторный аппарат улитки, содержащий рецепторные волосковые клетки (вторично-чувствующие механорецепторы). Эти клетки трансформируют механические колебания в электрические потенциалы.
Передача звуковых колебаний по каналам улитки происходит в результате того, что мембрана овального окна преддверия прогибается под влиянием силы звуковой волны, передавшейся через зукопроводящую часть уха. Это вызывает колебания перилимфы в верхнем и нижнем каналах улитки, которые доходят до круглого окна улитки. Преддверная мембрана очень тонкая, поэтому жидкость в верхнем и среднем каналах колеблется так, как будто оба канала едины. Упругим элементом, отделяющим этот как бы общий верхний канал от нижнего, является основная мембрана. Колебания, распространяющиеся по перилимфе и эндолимфе верхнего и среднего каналов как бегущая волна, приводят в движение эту мембрану и через нее передаются на перилимфу нижнего канала.
На основной мембране расположены два вида рецепторных волосковых клеток (вторично-чувствующих механорецепторов): внутренние (7) и наружные(8) отделенные друг от друга кортиевыми дугами (17). Внутренние волосковые клетки располагаются в один ряд; общее число их по всей длине перепончатого канала достигает 3500. Наружные волосковые клетки располагаются в 3-4 ряда, общее число их 12000-20000. Каждая волосковая клетка имеет удлиненную форму; один ее полюс фиксирован на основной мембране, второй находится в полости перепончатого канала улитки. На конце этого полюса есть волоски, или стереоцилии. Волоски рецепторных клеток омываются эндолимфой и контактируют с покровной (текториальной) мембраной (9), которая по всему ходу перепончатого канала расположена над волосковыми клетками.
Механизм восприятия звука. Жесткость основной мембраны быстро уменьшается в направлении от основания к верхушке улитки, а масса на единицу длины изменяется наоборот, т. е. увеличивается. Такая неоднородность механических свойств имеет существенное значение для распределения по длине улитки амплитуд колебаний, возникших в ответ на звуковой стимул.
По данным Бекеши, текториальная и основная мембраны колеблются синфазно, но имеют различные точки закрепления, одновременное смещение обеих мембран вызывает изгиб волосковых рецепторов и тем самым их возбуждение. Следовательно, характер движения основной мембраны определяет степень возбуждения волосковых клеток.
Рассмотрим процесс формирования колебаний основной мембраны несколько подробнее.
Под действием внешней силы мембрана овального окна прогибается и вызывает смещение соответствующего объема перилимфы. При этом ввиду несжимаемости перилимфы, должна выгнуться на эквивалентную величину мембрана круглого окна. При медленном смещении стремечка частицы перилимфы перемещаются вдоль основной мембраны по вестибулярному каналу и затем через геликотрему и по барабанному каналу к круглому окну. Пусть теперь на улитку действует быстрый стимул, например скачок звукового давления, вызывающий внезапное смещение мембраны овального окна на постоянную величину. Частицы жидкости, сдвинутые смещением мембраны овального окна, создают избыточное давление в вестибулярном канале улитки. Под действием этого давления быстрее всего смещается в сторону барабанной лестницы наименее инерционный базальный участок основной мембраны. В силу несжимаемости жидкости эквивалентный объем перилимфы выгибает мембрану круглого окна так, чтобы общий объем, занимаемый перилимфой в каналах улитки, остался неизменным. Однако в силу своей упругости смещенный участок основной мембраны стремится вернуться в исходное состояние и при этом сдвинуть мешающие ему частицы перилимфы. Последние перемещаются в сторону менее упругих и более инерционных участков основной мембраны, заставляя их в свою очередь прогибаться в сторону барабанной лестницы. Обратный процесс будет происходить в барабанной лестнице: выгиб мембраны круглого окна вызовет поток частиц перилимфы с общим направлением от геликотремы к базальной части барабанной лестницы. Деформация основной мембраны при своем распространении к геликотреме постепенно затухает, и практически отражения волны от конца улитковой перегородки не происходит. Таким образом, в ответ на быстрое смещение стремечка на основной мембране возникает колебательная реакция типа бегущей волны. Описанные выше представления о характере колебаний основной мембраны составляют основу концепции бегущих волн, сформулированной Бекеши в результате многолетних фундаментальных исследований улитки (Bekesy, 1960). Методом прямых наблюдений колебаний перегородки улитки под воздействием звуковых стимулов Бекеши смог измерить амплитуду колебаний различных точек основной мембраны при тональных сигналах нескольких частот (50, 100, 200 и 300 гц). Кроме того, для тех же тонов были определены сдвиги фаз между колебаниями стремечка и рассматриваемых точек на основной мембране. Совокупность полученных экспериментальных данных позволила установить факт возникновения бегущей волны по перегородке в ответ на гармонические колебания стремечка. Кучность гидродинамической волны, падающей на базальную мембрану, зависит от частоты звуковых волн. Колебания овального окна с определенной частотой вызывают бегущую волну, амплитуда которой возрастает до определенной точки на основной мембране, характерной для данной частоты. После достижения точки максимума бегущая волна быстро затухает при своем продвижении к геликотреме. Чем ниже частота возбуждающего сигнала, тем ближе к геликотреме находится точка максимального отклика. Нормированные амплитудно-координатные и фазо-координатные характеристики колебания улитки, экспериментально получены Бекеши с помощью измерений на препаратах органа слуха умерших людей. Скорость распространения волны в начале мембраны приблизительно в 10 раз больше, чем в районе геликотремы. Следовательно, и длина волны колебаний при их распространении по основной мембране должна резко уменьшаться к концу улитки. Эффект уменьшения длины волны колебаний, бегущих по основной мембране, при их приближении к геликотреме выражен отчетливо. Таким образом, даже при возбуждении улитки стационарным гармоническим сигналом колебания мембраны наряду со стационарной картиной распределения амплитуд смещений различных точек должны характеризоваться и динамическим процессом непрерывного изменения во времени фаз колебаний точек основной мембраны вдоль улитки. По-видимому, оба указанных фактора имеют значение при возбуждении рецепторной системы в кортиевом органе. При действии звука основная мембрана начинает колебаться, наиболее длинные волоски рецепторных клеток (стереоцилии) касаются покровной мембраны и несколько наклоняются. Отклонение волоска на несколько градусов приводит к натяжению тончайших вертикальных нитей (микрофиламент), связывающих между собой верхушки соседних волосков данной клетки. Это натяжение чисто механически открывает от 1 до 5 ионных каналов в мембране стереоцилии. Через открытый канал в волосок начинает течь калиевый ионный ток. Сила натяжения нити, необходимая для открывания одного канала, ничтожна, около 2•10-13ньютонов. Еще более удивительным кажется то, что наиболее слабые из ощущаемых человеком звуков растягивают вертикальные нити, связывающие верхушки соседних стереоцилии, на расстояние, вдвое меньшее, чем диаметр атома водорода.
Тот факт, что электрический ответ слухового рецептора достигает максимума уже через 100-500 мкс (микросекунд), означает, что ионные каналы мембраны открываются непосредственно механическим стимулом без участия вторичных внутриклеточных посредников. Это отличает механорецепторы от значительно медленнее работающих фоторецепторов.
Деполяризация пресинаптического окончания волосковой клетки приводит к выходу в синаптическую щель нейромедиатора (глутамата или аспартата). Воздействуя на постсинаптическую мембрану афферентного волокна, медиатор вызывает генерацию в нем возбуждающего постсинаптического потенциала и далее генерацию распространяющихся в нервные центры импульсов.
Открывания всего нескольких ионных каналов в мембране одной стереоцилии явно мало для возникновения рецепторного потенциала достаточной величины. Важным механизмом усиления сенсорного сигнала на рецепторном уровне слуховой системы является механическое взаимодействие всех стереоцилии (около 100) каждой волосковой клетки. Оказалось, что все стереоцилии одного рецептора связаны между собой в пучок тонкими поперечными нитями. Поэтому, когда сгибаются один или несколько более длинных волосков, они тянут за собой все остальные волоски. В результате этого открываются ионные каналы всех волосков, обеспечивая достаточную величину рецепторного потенциала.
Электрические явления в улитке. При отведении электрических потенциалов от разных частей улитки обнаружено пять различных феноменов: два из них — мембранный потенциал слуховой рецепторной клетки и потенциал эндолимфы — не обусловлены действием звука; три электрических явления — микрофонный потенциал улитки, суммационный потенциал и потенциалы слухового нерва — возникают под влиянием звуковых раздражений. Если ввести в улитку электроды, соединить их с динамиком через усилитель и подействовать на ухо звуком, то динамик точно воспроизведет этот звук. Описываемое явление называют микрофонным эффектом улитки, а регистрируемый электрический потенциал назван кохлеарным микрофонным потенциалом. Доказано, что он генерируется на мембране волосковой клетки в результате деформации волосков. Частота микрофонных потенциалов соответствует частоте звуковых колебаний, а амплитуда потенциалов в определенных границах пропорциональна интенсивности звука.
В ответ на сильные звуки большой частоты (высокие тона) отмечают стойкий сдвиг исходной разности потенциалов. Это явление получило название суммационного потенциала. Различают положительный и отрицательный суммационные потенциалы. Их величины пропорциональны интенсивности звукового давления и силе прижатия волосков рецепторных клеток к покровной мембране.
Микрофонный и суммационный потенциалы рассматривают как суммарные рецепторные потенциалы волосковых клеток. Имеются указания, что отрицательный суммационный потенциал генерируется внутренними, а микрофонный и положительный суммационные потенциалы — наружными волосковыми клетками. И, наконец, в результате возбуждения рецепторов происходит генерация импульсного сигнала в волокнах слухового нерва.
Сигналы от волосковых клеток поступают в мозг по афферентным нервным волокнам VIII пары черепных нервов. Они являются дендритами ганглиозных нервных клеток спирального ганглия. Около 90 % волокон идет от внутренних волосковых клеток и лишь 10% — от наружных. Сигналы от каждой внутренней волосковой клетки поступают в несколько волокон, в то время как сигналы от нескольких наружных волосковых клеток конвергируют на одном волокне.
Дата добавления: 2015-12-11; просмотров: 1318;