У рефлекторной реакции может быть гормональное звено, что
характерно для регуляции функций внутренних органов - вегетативных функций в отличие от соматических функций, рефлекторная регуляция которых осуществляется только нервным путем (деятельность опорно-двигательного аппарата). Гормональное звено включается за счет дополнительной выработки биологически активных веществ. Например, при действии на экстерорецепторы сильных раздражителей (холод, жара, болевой раздражитель) возникает мощный поток афферентных импульсов, поступающих в ЦНС, при этом в кровь выбрасывается дополнительное количество адреналина мозговым слоем надпочечников и гормонов коры надпочечников, играющих адаптивную (защитную) роль.
А. Понятия.Гормоны - биологически активные вещества, вырабатываемые эндокринными железами или специализированными клетками, находящимися в различных органах (например, в поджелудочной железе, пищеварительном тракте).
По месту действия на органы-мишени или на другие эндокринные железы гормоны делят на две группы:
1. Эффекторные гормоны, действующие на клетки-эффекторы (например, инсулин, регулирующий обмен веществ в организме, повышает синтез гликогена в клетках печени, увеличивает транспорт глюкозы и других веществ через клеточную мембрану, повышает интенсивность синтеза белка).
2. Тропные гормоны, действующие на другие эндокринные железы и регулирующие их функции (например, адренокортикотропный гормон гипофиза - АКТГ регулирует выработку гормонов корой надпочечников).
Б. Виды влияния гормонов.Гормоны оказывают два вида влияния на органы, ткани и системы организма: функциональное (играют весьма важную роль в регуляции функций организма) и морфогенетическое - обеспечивают морфогенез (рост, физическое, половое и умственное развитие. Например, при недостатке тироксина страдает развитие ЦНС, следовательно, и умственное развитие).
1. Функциональное влияние гормонов бывает трех видов: пусковое, модулирующее и пермиссивное.
1)Пусковое влияние - это способность гормона запускать деятельность эффектора. Например, адреналин запускает распад гликогена в печени и выход глюкозы в кровь; вазопрессин (антидиуретический гормон - АДГ) включает реабсорбцию воды из собирательных трубок нефрона в интерстнций почки.
2) Модулирующее влияние гормона - изменение интенсивности протекания биохимических процессов в органах и тканях. Например, активация окислительных процессов тироксином, которые могут происходить и без него; стимуляция деятельности сердца адреналином, которая возможна и без адреналина. Модулирующим влиянием гормонов является также изменение чувствительности ткани к действию других гормонов. Например, фолликулин усиливает действие прогестерона на слизистую оболочку матки, тиреоидные гормоны усиливают эффекты катехоламинов.
3) Пермиссивное влияние гормонов - способность одного гормона обеспечивать реализацию эффекта другого гормона. Например, инсулин необходим для проявления действия соматотропного гормона, фоллитропин необходим для реализации эффекта лютропина.
2. Морфогенетическое влияние гормонов (рост, физическое и половое развитие) подробно изучается другими дисциплинами (гистология, биохимия) и лишь частично - в курсе физиологии. Оба вида влияния гормонов (морфогеиетическое и функциональное) реализуются с помощью метаболических процессов, запускаемых посредством клеточных ферментных систем.
В. Механизм действия гормонов. Если клеточная мембрана непроницаема для гормона, то эффекторные рецепторы расположены на ее поверхности (белково-полипептидные и аминокислотные гормоны гипоталамуса, гипофиза, поджелудочной железы, мозгового слоя надпочечников водорастворимы, но не растворимы в липидах). Если клеточная мембрана проницаема для гормона, то эффекторные рецепторы для этих гормонов расположены в основном в цитоплазме (стероидные гормоны - кортикоиды и половые гормоны растворимы в липидах). Гормоны щитовидной железы могут проникать в клетку только в комплексе со специальным белком плазмы крови, посредством эндоцитоза. Их рецепторы расположены как на поверхности клеточной мембраны, так и в цитоплазме.
Гормонрецепторный комплекс белково-пептидных гормонов и производных аминокислот активирует мембранные ферменты, что ведет к образованию вторых посредников, реализующих свое дей-
ствие в цитоплазме и ядре клетки. Ими являются: I) аденилатциклаза - циклический аденозинмонфосфат (цАМФ); 2) гуанилатциклаза - циклический гуанозинмонофосфат (цГМФ); 3) фосфолипаза С - инозитолтрифосфат (ИФз): 4) ионизированный кальций - кальмодулин (белок немышечных клеток) или тропонин С - в мышечных клетках. Гормонрецепторный комплекс стероидных гормонов и гормонов щитовидной железы, проникающих в клетку, соединяется в клетке с эффекторным рецептором - белком. Этот комплекс поступает в клеточное ядро, где происходит соединение с ядерным рецептором, после чего реализуется влияние данного гормона на клетку. Часть эффектов стероидных гормонов, как и гормонов щитовидной железы, реализуется посредством рецепторов, расположенных на клеточной мембране и вторых посредников, как и у белково-пептидных гормонов.
Действие гормонов прекращается с помощью тканевых ферментов и ферментов самих эндокринных желез, печени, почек. Многие продукты расщепления гормонов также активны и вызывают иногда сходные эффекты. Продукты распада гормонов выводятся главным образом почками, а также слюнными и потовыми железами, железами пищеварительного тракта и с желчью.
Г. Регуляция выработки гормонов осуществляется непосредственно нервной системой, но главным образом с помощью гормонов гипофиза, функция которого регулируется в свою очередь гормонами гипоталамуса - нейрогормонами (схема 1.1). Для некоторых эндокринных желез основным механизмом регуляции является местная саморегуляция. Так, секреция инсулина и глюкагона панкреатическими островками (островки Лангерганса) регулируется уровнем глюкозы в крови. Если концентрация глюкозы в крови высокая, то по принципу обратной отрицательной связи стимулируется выработка инсулина, который снижает концентрацию глюкозы в крови с помощью увеличения утилизации ее клетками организма и увеличения отложения в виде гликогена в клетках печени, в результате чего снижается (нормализуется) концентрация глюкозы в крови. В случае снижения концентрации глюкозы в крови выработка инсулина уменьшается, выработка глюкагона клетками островков Лангерганса возрастает (глюкагон увеличивает преобразование гликогена печени в глюкозу и выход ее в кровь). Секреция кальцийрегулирующих гормонов (парати-рина и кальцитонина) также регулируется по принципу обратной отрицательной связи - за счет концентрации кальция в крови.
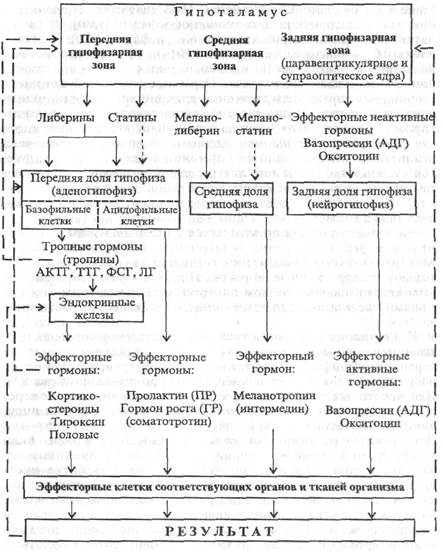
| Схема 1.1. Механизм гипоталамо-гипофизарных взаимодействий в регуляции выработки гормонов. АКТГ - адренокортикотропный гормон (кортикотропин), ТТГ - тнреотропный гормон (тиреотропин), ФС Г - фолликулостимулирующий гормон (фоллитропин), ЛГ-лютеинизирукшшй гормон (люгропин), АДГ - аптидиурстический гормон. Примечание. Сплошные стрелки - стимулирующее влияние, пунктирные - угнетающее. 1.3. РЕГУЛЯЦИЯ С ПОМОЩЬЮ МЕТАБОЛИТОВ И ТКАНЕВЫХ ГОРМОНОВ. МИОГЕННЫЙ МЕХАНИЗМ РЕГУЛЯЦИИ А. Метаболиты - продукты, образующиеся в организме в процессе обмена веществ как результат различных биохимических реакций. Это аминокислоты, нуклеотиды, коферменты, угольная, молочная, пировиноградная, адениловая кислоты, ионный сдвиг, что сопровождается изменением рН. |
Регуляция с помощью метаболитов на ранних этапах филогенеза была единственной.Метаболиты одной клетки непосредственно влияли на другую - соседнюю - клетку или группу клеток, которые в свою очередь таким же способом действовали на следующие клетки (контактная регуляция). С появлением гемолимфы и сосудистой системы метаболиты с движущейся гемолимфой стали передвигаться к другим клеткам организма, даже расположенным на большом расстоянии, причем процесс стал осуществляться быстрее. Затем появилась нервная система как регулирующая система, а еще позже - эндокринные железы. Метаболиты действуют в основном местно, но могут влиять и на другие органы и ткани, на активность нервных центров. Например, накопление угольной кислоты в крови ведет к возбуждению дыхательного центра и усилению дыхания. Примером местной гуморальной регуляции может служить гиперемия интенсивно работающей скелетной мышцы: накапливающиеся метаболиты обеспечивают расширение кровеносных сосудов, что увеличивает доставку кислорода и питательных веществ к мышце. Подобные регуляторные влияния метаболитов происходят и в других активно работающих органах и тканях организма.
Б. Тканевые гормоны: биогенные амины (гистамин, серотонин), простагландины и кинины.Они занимают промежуточное положение между гормонами и метаболитами как гуморальными веществами регуляции. Эти вещества свое регулирующее влияние оказывают на клетки тканей посредством изменения их биофизических свойств (проницаемость мембран, их возбудимость), интенсивности обменных процессов, чувствительности клеточных рецепторов, образования вторых посредников. В результате влияния тканевых гормонов меняется чувствительность клеток к нервным и гуморальным влияниям. Поэтому тканевые гормоны называют модуляторами регуляторных сигналов - они оказывают модулирующее влияние. Тканевые гормоны образуются неспециализированными клетками, но действуют посредством специализированных клеточных рецепторов: например, для гистамина обнаружено два вида рецепторов - Н] и Н2. Известно, что тканевые гормоны влияют напроницаемость клеточных мембран: они регулируют поступление в клетку и выход из нее различных веществ и ионов, определяющих мембранный потенциал, а значит, и ПД.
В. Многснный механизм регуляции.С развитием мышечной системы в процессе эволюции миогенный механизм регуляции функций постепенно становится все более заметным. Организм человека по своей массе примерно на 50% состоит из мышц. Это скелетная мускулатура (40% от массы тела), мышца сердца, гладкие мышцы кровеносных и лимфатических сосудов, стенки пищеварительного тракта, желчного и мочевого пузырей и других внутренних органов. Сущность миогенного механизма регуляции состоит в том, что предварительное умеренное растяжение скелетной или сердечной мышцы увеличивает силу их сокращений. Сократительная активность гладкой мышцы также зависит от степени наполнения полого мышечного органа, а значит, и его растяжения. При увеличении наполнения органа тонус гладкой мышцы сначала возрастает, а затем возвращается к исходному уровню (пластичность гладкой мышцы), что обеспечивает регуляцию тонуса сосудов и наполнение внутренних полых органов без существенного повышения давления в них (до определенной величины). Кроме того, большинство гладких мышц обладает автома-тией, они постоянно находятся в некоторой степени сокращения под влиянием импульсов, возникающих в них самих (например, мышцы кишечника, кровеносных сосудов). Импульсы, поступающие к ним по вегетативным нервам, оказывают модулирующее влияние - увеличивают или уменьшают тонус гладких мышечных волокон.
Дата добавления: 2015-10-22; просмотров: 1156;