Сенсорные рецепторы
А. Понятия. Различают сенсорные и эффекторные рецепторы. Сенсорные рецепторы воспринимают изменения внешней или внутренней среды организма и обеспечивают поступление информации в ЦНС. Эффекторные рецепторы рабочих органов - эффекторов (сердце, скелетная мышца, желудок и т.д.) воспринимают сигналы (команды) от нервной системы в виде нервных импульсов, они реагируют также на гуморальные вещества, циркулирующие в крови (гормоны, медиаторы, метаболиты). Нервные клетки передают сигналы друг другу также посредством рецепторов, которые тоже являются эффекторными. В данном разделе рассматриваются только сенсорные рецепторы.
Рецепторы (от лат. receptum - принимать) приспособлены воспринимать обычно какой-то один вид раздражителей (адекватный) и максимально чувствительны к нему. Например, рецепторы сетчатки реагируют на свет, но не возбуждаются при воздействии звукового раздражителя. К другим - неадекватным раздражителям -они малочувствительны. Однако неадекватные раздражители такжемогут возбудить сенсорные рецепторы. Например, механическое воздействие на глаз вызывает ощущение света, однако энергия неадекватного раздражителя должна быть во много раз больше энергии адекватного.
Б. Классификация сенсорных рецепторов. Нервная система отличается большим разнообразием рецепторов (рис. 1.8).
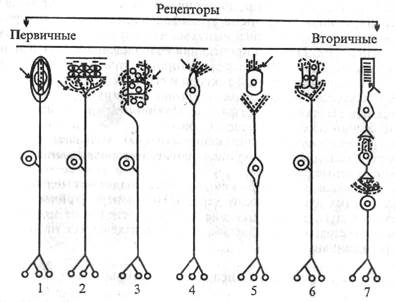
| Рис. 1.8. Различные типы рсцепторных клеток. Стрелки указывают на участки действия раздражителей (по Г. Шсперду, 1987, с изменениями). 1,2- соматосенсорные рецепторы; 3 -• мышечный рецептор; 4 - обонятельный рецептор; 5 - слуховой и вестибулярный рецепторы; 6 - вкусовой рецептор; 7 - зрительный рецептор 1. Согласно структурно-функциональной организации, различают первичные и вторичные рецепторы. Первичные рецепторы представляют собой чувствительные окончания дендрита сенсорного нейрона, тело нейрона расположено в спинномозговом ганглии или в ганглиях черепных нервов. В первичном рецепторе |
раздражитель действует непосредственно на окончания дендрита сенсорного нейрона.
Вторичные рецепторы имеют специальную клетку, синаптиче-ски связанную с окончанием дендрита сенсорного нейрона. Ко вторичным рецепторам относятся фоторецепторы, слуховые, вестибулярные, вкусовые рецепторы.
2. По скорости адаптации рецепторы делят на три группы: быстро адаптирующиеся (фазные), медленно адаптирующиеся (тонические) и смешанные (фазнотонические), адаптирующиеся со средней скоростью. Примером быстро адаптирующихся рецепторов являются кожные рецепторы вибрации (тельца Пачини) и прикосновения (тельца Мейснера). К медленно адаптирующимся рецепторам относятся проприорецепторы, рецепторы растяжения легких, болевые рецепторы. Со средней скоростью адаптируются фоторецепторы сетчатки, терморецепторы кожи.
3. В зависимости от вида воспринимаемого раздражителя выделяют 5 типов рецепторов, 1) Механореиетпоры возбуждаются при механической их деформации. Они расположены в коже, внутренних органах, опорно-двигательном аппарате, слуховой и вестибулярной системах. 2) Хеморецепторы воспринимают химические изменения внешней и внутренней среды организма. Такие рецепторы есть в слизистой оболочке языка (вкусовые рецепторы), носа (обонятельные рецепторы), каротидном и аортальном тельцах, продолговатом мозге, гипоталамусе. 3) Терморецепторы воспринимают изменения температуры. Они подразделяются на тепловые и холодовые рецепторы, находятся в коже, сосудах, внутренних органах, в продолговатом и спинном мозге, в среднем мозге и гипоталамусе. 4) Фоторецеппюры воспринимают световую (электромагнитную) энергию, расположены в сетчатке глаза. 5) Ноцицепторы (болевые рецепторы) отвечают на механические, термические, химические (тетании, брадикинин, К+. Н* и др.) раздражители. Болевые стимулы воспринимаются свободными нервными окончаниями.
4. По расположению в организме выделяют экстеро- и интеро-рецепторы. К экстерорецепторам относятся рецепторы кожи, видимых слизистых оболочек и органов чувств. Интерореиепторами являются рецепторы внутренних органов (висцерорецепторы), сосудов и ЦНС; проприорецепторы - рецепторы опорно-двигательного аппарата и вестибулярные рецепторы.
5. С психофизиологической точки зрения рецепторы подразделяются в соответствии с органами чувств и формируемыми ощущениями на зрительные, слуховые, тактильные, вкусовые и обонятельные.
6. В зависимости от степени специфичности рецепторов, т.е. способности отвечать на один или более видов раздражителей, выделяют мономодальные рецепторы, воспринимающие один вид раздражителей и полимодальные, воспринимающие два-три вида раздражителей. Мономодальными рецепторами являются зрительные, вкусовые, обонятельные. Различия в чувствительности к адекватным и неадекватным раздражителям у полимодальных рецепторов выражены меньше, чем у мономодальных. Пример полимодальных рецепторов - болевые рецепторы.
В. Свойства рецепторов. 1.Высокая возбудимость рецепторов. Например, волосковые рецепторы внутреннего уха способны обнаружить движение мембраны кортиева органа, равное диаметру атома водорода; для возбуждения фоторецептора сетчатки достаточно одного кванта света, обонятельного рецептора - одной молекулы пахучего вещества. Возбудимость различных рецепторов неодинакова. У висцерорецепторов она ниже, чем у экстерорецепторов. Высокая возбудимость рецепторов обеспечивает восприятие малейших изменений внутренней и внешней среды, что необходимо для надежной регуляции функций внутренних органов и приспособления организма в окружающей среде.
2. Спонтанная активность рецепторов. Многие виды рецепторов (фото-, фоно-, вестибуло-, термо-, хемо- и проприорецепторы) генерируют в окончании сенсорного нейрона потенциалы действия (ПД) в отсутствие раздражителя. Это связано со спонтанным колебанием мембранного потенциала в рецепторе, которое периодически достигает критического уровня деполяризации, что приводит к генерации ПД в нервном волокне. Возбудимость таких рецепторов выше, чем возбудимость рецепторов без фоновой активности. Значение фоновой активности рецепторовзаключается в том, что она участвует в поддержании тонуса нервных центров в условиях физиологического покоя и бодрствующего состояния организма.
3. Адаптация рецепторов - уменьшение их возбудимости при длительном действии раздражителя. Процессы адаптации, формирующиеся на разных этапах преобразования информации, приводят к снижению амплитуды рецепторного потенциала и, как следствие, уменьшается частота импульсации сенсорного нейрона. Механизм адаптации рецепторов изучен недостаточно. Одним из факторов является накопление Са2+ внутри клетки при ее возбуждении, что может ингибировать каскад ферментативных реакций в рецепторе. Другим возможным механизмом действия Са2+ является активирование Са-зависимых калиевых каналов. Выход К+ через эти каналы из клетки препятствует деполяризации ее мембраны при формировании рецепторного потенциала.
Значение адаптациирецепторов заключается в том, что предотвращается избыточное поступление информации в ЦНС и устраняются неприятные ощущения.
Г. Механизм восприятия раздражителя, действующего на рецепторы.Несмотря на большое многообразие рецепторов, в каждом из них можно выделить два или три этапа преобразования энергии раздражения в нервный импульс в зависимости от структурной организации рецептора. Первый этап - возникновение рецепторного потенциала при действии раздражителя на рецептор - одинаковый для всех рецепторов.
Действие раздражителя способно вызывать конформационные изменения рецепторного белка. На данном этапе во многих рецепторах происходит усиление сигнала, так как энергия формирующегося рецепторного потенциала оказывается многократно (например, в фоторецепторе в 105 раз) больше пороговой энергии раздражения.
В рецепторах (кроме фоторецепторов) энергия раздражителя приводит к открытию натриевых каналов и появлению ионных токов, среди которых основную роль играет входящий натриевый ток. Он приводит к деполяризации мембраны рецептора. Считают, что в хеморецепторах открытие каналов связано с изменением формы (конформацией) белковых молекул, выполняющих функцию ворот, а в механореценторах - с растяжением мембраны и расширением каналов. В фоторецепторах ионы натрия проникают в темноте в клетку, при действии света натриевые каналы закрываются, что уменьшает входящий натриевый ток. В связи с этим рецепторный потенциал представлен не деполяризацией, а гиперполяризацией вследствие выхода К* из клетки при уменьшении входа На+ в клетку.
Чтобы информация сенсорных раздражителей достигла ЦНС, рецепторный потенциал должен быть преобразован в ПД. В первичных и вторичных рецепторах это происходит разными способами. Второй этап в первичных и вторичных рецепторах протекает по-разному. В первичном рецепторе рецеп-торная зона является частью окончаний сенсорного нейрона. Возникший рецепторный потенциал вызывает деполяризацию в участке афферентного волокна, в котором возможно возникновение ПД. В миелиновых волокнах ПД возникает в ближайших перехватах Ранвье, в безмиелиновых - в ближайших участках, имеющих достаточную концентрацию потенциалзави-симых натриевых и калиевых каналов. Если деполяризация мембраны при этом достигает критического уровня, происходит генерация ПД (рис. 1.9), т.е. процесс возникновения возбуждения происходит в два этапа.
Во вторичных рецепторахпреобразование энергии раздражения в нервный импульс происходит в три этапа. На I этапе возникает рецепторный потенциал в эпителиальной рецепторной клетке, синаптически связанной с окончанием сенсорного нейрона. Рецепторный потенциал вызывает выделение в синаптическую щель медиатора. Под влиянием медиатора на постсинаптической мембране возникает генераторный потенциал (возбуждающий постсинаптический потенциал) - это II этап. Ш этап - возникновение ПД в нервном волокне вблизи постсинаптической мембраны под влиянием генераторного потенциала.
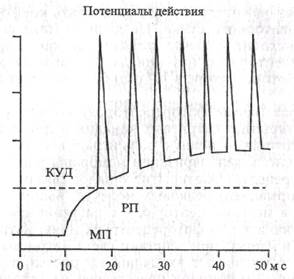
| Рис. 1.9. Типичные взаимоотношения между рецепторным потенциалом и потенциалом действия при сверхпороговом уровне рецепторного потенциала (по А. Гайтону, 1986, с изменениями). КУД - критический уровень деполяризации; РП - рецепторный потенциал; МП - мембранный потенциал После поступления афферентных импульсов от рецепторов (рефлексогенных зон) в нервный центр формируется ответная рефлекторная реакция организма на раздражения. Основные положения рефлекторного принципа регуляции сформировались на протяжении весьма длительного отрезка времени. 1.1.7. Развитие концепции рефлекса. Нервизм и нервный центр А. Основные положения рефлекторного принципа деятельности ЦНС были разработаны на протяже- |
нии примерно двух с половиной веков. Можно выделить пять основных этапов развития этой концепции.
Этап I - заложены основы понимания рефлекторного принципа деятельности ЦНС французским естествоиспытателем и математиком Р1Декартом (XVII в.). Р.Декарт считал, что «все вещи и явления можно объяснить естественнонаучным путем». Эта исходная позиция позволила Р.Декарту сформулировать два важных положения рефлекторной теории:
1) всякая реакция на внешнее воздействие является отраженной (впоследствии ее стали называть рефлекторной);
2) ответная реакция на раздражение осуществляется при помощи нервной системы. Однако Р. Декарт был дуалистом.
По Р.Декарту, нервы - это трубочки, по которым с огромной скоростью движутся животные духи, материальные частицы неизвестной природы, по нервам они попадают в мышцу и мышца раздувается (сокращается).
Этап II - экспериментальное обоснование материалистических представлений о рефлексе (ХVII-Х1Х вв.). В частности, было установлено, что рефлекторная реакция может осуществляться на одном метамере лягушки (сегмент спинного мозга, связанный с «кусочком тела»). Выявлено, что стимулы могут быть не только внешними, но и внутренними, установлена роль задних - чувствительных и передних — двигательных корешков спинного мозга (закон Белла - Мажанди).
Весьма активно сегментарные рефлексы изучал Ч.Шеррингтон (конец XVIII- начало XIX в.).
Этап III - победа материалистических представлений о психической деятельности (И.М.Сеченов, 60-е гг. XIX в.). Наблюдая за развитием детей, И. М. Сеченов пришел к заключению, что в основе формирования психической деятельности лежит принцип рефлекса. Это положение он выразил следующей фразой: «Все акты сознательной и бессознательной жизни по способу происхождения суть рефлексы». Таким образом, И.М.Сеченов стал на путь детерминизма в вопросах психической деятельности человека. При изучении рефлексов он обосновал приспособительный характер изменчивости рефлекса, открыл торможение рефлексов (центральное торможение; 1863), суммацию возбуждения в ЦНС (1868).
Этап IV - разработаны основы учения о высшей нервной деятельности (И.П.Павлов, начало XX в.). И.П.Павлов открыл условные рефлексы и использовал их как объективный метод изучения психической деятельности (высшей нервной деятельности — по И.П.Павлову). Он сформулировал 3 принципа рефлекторной теории:
1. Принцип детерминизма (принцип причинности), согласно которому любая рефлекторная реакция причинно обусловлена.
2. Принцип структурности, суть которого заключается в том, что каждая рефлекторная реакция осуществляется с помощью определенных структур и чем больше структурных элементов участвует в осуществлении этой реакции, тем она совершеннее.
3. Принцип единства процессов анализа и синтеза в составе рефлекторной реакции: нервная система анализирует (различает) с помощью рецепторов все действующие внешние и внутренние раздражители и на основании этого анализа формирует целостную ответную реакцию (синтез).
Этап V - создано учение о функциональных системах (П.К.Анохин, середина XXв.; см. раздел 1.5).
Б. Нервизм- это концепция, признающая ведущую роль нервной системы в регуляции функций всех органов и тканей организма (физиологическийнервизм). Концепция нервизма прошла весьма длинный исторический путь своего развития. Почву для этой концепции подготовил Р.Декарт (1596-1650), выдвинувший идею о рефлекторном принципе деятельности нервной системы. Ф.Гофман (1660-1742) сформулировал гипотезу о влиянии нервов на «все перемены в здоровом и больном состоянии». Согласно У.Кулену (1712-1790), все процессы в здоровом и больном организме регулирует «нервный принцип», который проявляет свое действие через головной мозг при посредстве нервов - проводников нервной деятельности. По мнению Е.О.Мухина (1817), «все человеческое тело вообще можно, отвлекаясь, рассматривать как построенное из нервов, ибо остальные части тела, видимо, существуют вследствие нервов, как управляющих их способностями».
Велики заслуги в развитии концепции нервизма И.М.Сеченова (1829-1905) и "С.П.Боткина (1832-1889). По мнению С.П.Боткина, организм - это целостная система, деятельность которой направляется и регулируется нервной системой. Он рассматривал различные заболевания как следствие нарушения нормальных нервных соотношений - клинический нервизм.Нарушение «регуляторных нервных аппаратов» может явиться причиной ряда болезней человека, что убедительно подтверждено клиническими наблюдениями.
Выдающийся вклад в развитие концепции физиологического нервизма сделал И.П.Павлов (1849-1936). Он обосновал представление о трофическом влиянии нервной системы на органы и ткани, сформулировал принципы рефлекторной теории, доказал важную роль нервной системы в регуляции секреции желез пищеварительного тракта, открыл условные рефлексы и с их помощью разработал основы учения о высшей нервной деятельности.
В. Нервный центр- это совокупность нейронов, расположенных на различных уровнях ЦНС, достаточных для приспособительной регуляции функции органа согласно потребностям организма. Например, нейроны дыхательного центра локализуются и в спинном мозге, и в продолговатом мозге, и в мосту. Однако среди нескольких групп клеток, расположенных на различных уровнях ЦНС, обычно имеется главная часть центра. Главная часть дыхательного центра находится в продолговатом мозге и включает инспираторные и экспираторные нейроны.
Нервный центр реализует свое влияние на эффекторы либо непосредственно с помощью эфферентных импульсов соматической и вегетативной нервных систем, либо с помощью активации вы-
работки соответствующих гормонов. Кроме нервной регуляции, в организме существуют гуморальный и миогенный механизмы регуляции. Гуморальный механизм регуляции функций органов и тканей организма осуществляется с помощью гормонов, медиаторов, метаболитов и тканевых гормонов.
Дата добавления: 2015-10-22; просмотров: 2188;