Локализация ферментов после электрофореза
В ряде случаев электрофорез ферментов в мягких условиях
(без ДДС-Na или мочевины) не сказывается на их активности.
Положение полос ферментов в геле можно идентифицировать с
помощью специфических реакций, дающих окрашенные продук-
ты. Для этого гель вымачивают в растворе субстратов и кофак-
торов, необходимых для протекания определенной ферментатив-
ной реакции, иногда с добавлением реагентов, сообщающих
окраску продукту этой реакции. Эту операцию следует проводить
быстро во избежание диффузии нефиксированных белков из их
полос. Если субстрат реакции полимерен и плохо диффундирует,
его можно заполимеризовать прямо в гель и следить за продви-
жением фронта ферментативной реакции вместе с миграцией
фермента. Разумеется, при этом необходимо ясно представлять,
каково будет поведение субстрата в электрическом поле во вре-
мя электрофореза.
Можно заполимеризовать субстрат и кофакторы в другом
(«индикаторном») геле на основе агарозы или импрегнировать
ими фильтровальную бумагу. После окончания электрофореза
белков в пластине рабочего ПААГ на его поверхность наклады-
вают индикаторный гель или бумагу. Ферменты диффундируют
в них из рабочего геля и обнаруживают себя соответствующей
цветной реакцией. Габриель привел длинный список индикатор-
ных цветных реакций для ферментов [Gabriel, 1971]. Во многих
случаях в этих реакциях фигурирует восстановление НАД с
дальнейшей передачей электрона на краситель. Удобно в каче-
стве заключительного этапа реакции воспользоваться нефермен-
тативным восстановлением тетразолиевой соли до ее формазана,.
что дает сильно окрашенные и зачастую нерастворимые продук-
ты (схема реакции приведена ниже),
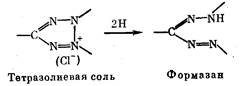
В качестве примера можно назвать часто употребляющийся
препарат «Methyl thiazolyl blue» (MTT) или еще более сложное
соединение «Nitro blue tetrazolium» (NBT). В окисленном виде
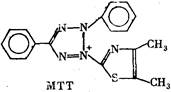
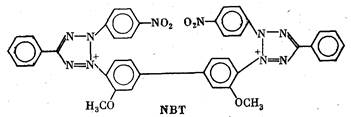
обе эти соли бесцветны. При восстановлении первая дает сине-
пурпурную, а вторая—иссиня-черную окраску. Реакция восста-
новления красителей ускоряется в присутствии промежуточного
переносчика электронов «Phenazine methosulfate» (PMS) или
нового, более стойкого реагента, выпускаемого для этой цели
фирмой «Boehringer» под названием «Meldolablue» [Turner,
Hopkinson, 1979].
Для обнаружения в геле протеолитических ферментов пред-
ложен ряд нетривиальных методов. Например, после электрофо-
реза гель вымачивают в течение 2 ч при 37° в 8%-ном растворе
цитохрома с в буфере с рН 8,5, содержащем 8 М мочевину и
1,7 мМ CaCl2. Затем белок фиксируют в 12,5%-ной ТХУ. Диф-
фундируя в гель, цитохром с окрашивает его в коричневый цвет
по всему объему. Полосы протеаз, разрушающих цитохром с,
остаются прозрачными и хорошо видны на коричневом фоне
[Ward, 1976]. Суть другого подхода заключается в том, что на
гель накладывают обработанную закрепителем в темноте (для
удаления серебра) и вымоченную в буфере фотопленку. После
трехчасовой выдержки при 35° во влажной камере на фотоплен-
ке можно обнаружить следы диффузии в нее протеаз, разрушаю-
щих слой желатины [Burger, Schroeder, 1976].
Для обнаружения нуклеаз при полимеризации геля в него
вносят ДНК (10 мкг/мл) или РНК (25 мкг/мл). Если нуклеазы
для своего действия нуждаются в ионах Mg2+, Са2+ или Zn2+, то
в их растворе ПААГ вымачивают после окончания электрофоре-
за. Далее следует окрашивание геля красителем для нуклеино-
вых кислот—бромистым этидием (см. главу 4). Окрашивается
весь гель, за исключением полос, содержащих нуклеазы, где
нуклеиновая кислота разрушена. Можно вести разделение нук-
леаз и в присутствии ДДС-Na, блокирующего их действие во
время электрофореза. Перед обработкой бромистым этидием
ДДС-Na из геля вымывают.
Заполимеризовав в ПААГ денатурированную ДНК, можно
выявить нуклеазы, специфичные для однонитевых ДНК. Мож-
но также исследовать оптимизацию условий, необходимых для
активности нуклеаз, варьируя состав буфера геля, или опробо-
вать различные ингибиторы нуклеазной активности. С помощью
кольцевых ДНК удается отличить эндонуклеазы от экзонуклеаз.
Электрофорезу можно подвергнуть лизаты бактерий для выяс-
нения содержания в них нуклеаз [Rosenthal, Lacks, 1977]. Окра-
шивание бромистым этидием в описанных опытах можно заме-
нить авторадиографией (см. ниже), если в гель заполимеризо-
вать меченную 32Р нуклеиновую кислоту или синтетический
полинуклеотид [Iborra et al., 1979].
Выше указывалось, что для локализации ферментов поих
активности электрофорез желательно проводить в условиях,
исключающих возможность денатурации. Однако в недавней ра-
боте [Lacks, Springhorn, 1980] было показано, что очистку цело-
го ряда ферментов (ДНКазы I, некоторых протеаз, амилазы,
глюкозо-6-фосфатдегидрогеназы) можно производить электро-
форезом в ПААГ с ДДС-Na после соответствующей обработки
их детергентом при повышенной температуре. Денатурация ока-
зывается обратимой. По окончании электрофореза ДДС-Na от-
мывают и одновременно снимают с белка встряхиванием геля в
течение 1—2 ч в 0,04 М Трис-буфере, рН 7,6. Ферментативная
активность в ряде случаев восстанавливается.
По данным Хагера и Буржесс, при снятии ДДС-Na с фермен-
та имеет смысл денатурировать его кратковременной обработ-
кой 6 М гуанидинхлоридом с последующей ренатурацей при раз-
бавлении. Возможно, что при этом восстанавливается нативная
конфигурация белка, нарушенная при первоначальной обработке
ДДС-Na [Hager, Burgess, 1980]. Для ферментов, содержащих
несколько полипептидных цепей (химотрипсин), предваритель-
ную обработку (b-меркаптоэтанолом, разумеется, следует исклю-
чить. Впрочем, то же самое относится и к одноцепочечному трип-
сину. По-видимому, разрыв внутрицепочечных S—S-связей в
нем приводит к необратимой денатурации. Ферменты, состоящие
более чем из двух субъединиц, не ренатурируют.
В заключение отметим, что гликопротеиды после электрофо-
реза в ПААГ можно локализовать, вымачивая гель в растворе
конканавалина А, конъюгированного с флюоресцентной меткой,
например с флюоресцеинизотиоцианатом [Furlan et al., 1979],
или радиоактивно меченного введением 125I [Horst et al., 1980].
Дата добавления: 2015-10-21; просмотров: 1200;