Восприятие высоты, силы звука и локализации источника звука
Восприятие высоты, силы звука и локализации источника звука начинается с попадания звуковых волн в наружное ухо, где они приводят в движение барабанную перепонку. Колебания барабанной перепонки через систему слуховых косточек среднего уха передаются на мембрану овального окна, что вызывает колебание перилимфы вестибулярной (верхней) лестницы. Эти колебания через геликотрему передаются перилимфе барабанной (нижней) лестницы и доходят до круглого окна, смещая его мембрану по направлению к полости среднего уха (рис. 2.8).
Колебания перилимфы передаются также на эндолимфу перепончатого (среднего) канала, что приводит в колебательные движения основную мембрану, состоящую из отдельных волокон, натянутых, как струны рояля. При действии звука волокна мембраны приходят в колебательные движения вместе с рецепторными клетками кортиева органа, расположенными на них. При этом волоски рецепторных клеток контактируют с текториальной мембраной, реснички волосковых клеток деформируются. Возникает вначале рецепторный потенциал, а затем потенциал действия (нервный импульс), который далее проводится по слуховому нерву и передается в другие отделы слухового анализатора.
Электрические явления в улитке.В улитке можно зарегистрировать пять различных электрических феноменов.
1. Мембранный потенциал слуховой рецепторной клетки характеризует состояние покоя.
2. Потенциал эндолимфы, или эндокохлеарный потенциал, обусловлен различным уровнем окислительно-восстановительных процессов в каналах улитки, в результате чего возникает разность потенциалов (80 мВ) между перилимфой среднего канала улитки (потенциал которой имеет положительный заряд) и содержимым верхнего и нижнего каналов. Этот эндокохлеарный потенциал оказывает влияние на мембранный потенциал слуховых рецепторных клеток, создавая у них критический уровень поляризации, при котором незначительное механическое воздействие во время контакта волосковых рецепторных клеток с текториальной мембраной приводит к возникновению в них возбуждения.
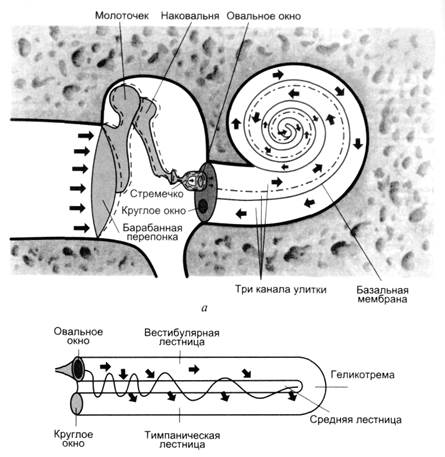
Рис. 2.8. Каналы улитки:
а – среднее и внутреннее ухо в разрезе (по П. Линдсею и Д. Норману, 1974); б – распространение звуковых колебаний в улитке
3. Микрофонный эффект улитки был получен в эксперименте на кошках. Электроды, введенные в улитку, соединялись с усилителем и громкоговорителем. Если рядом с ухом кошки произносили различные слова, то их можно услышать, находясь у громкоговорителя в другом помещении. Этот потенциал генерируется на мембране волосковой клетки в результате деформации волосков при соприкосновении с текториальной мембраной. Частота микрофонных потенциалов соответствует частоте звуковых колебаний, а амплитуда потенциалов в определенных границах пропорциональна интенсивности звуков речи. Звуковые колебания, действующие на внутреннее ухо, приводят к тому, что возникающий микрофонный эффект накладывается на эндокохлеарный потенциал и вызывает его модуляцию.
4. Суммационный потенциал отличается от микрофонного потенциала тем, что отражает не форму звуковой волны, а ее огибающую. Он представляет собой совокупность микрофонных потенциалов, возникающих при действии сильных звуков с частотой выше 4000 – 5000 Гц. Микрофонный и суммационный потенциалы связывают с деятельностью наружных волосковых клеток и рассматривают как рецепторные потенциалы.
5. Потенциал действия слухового нерва регистрируется в его волокнах, частота импульсов соответствует частоте звуковых волн, если она не превышает 1000 Гц. При действии более высоких тонов частота импульсов в нервных волокнах не возрастает, так как 1000 имп/с – это почти максимально возможная частота генерации импульсов в волокнах слухового нерва. Потенциал действия в нервных окончаниях регистрируется через 0,5 –1,0 мс после возникновения микрофонного эффекта, что свидетельствует о синаптической передаче возбуждения с волосковой клетки на волокно слухового нерва.
Восприятие звуков различной высоты(частоты), согласно резонансной теории Гельмгольца, обусловлено тем, что каждое волокно основной мембраны настроено на звук определенной частоты. Так, звуки низкой частоты воспринимаются длинными волнами основной мембраны, расположенными ближе к верхушке улитки, звуки высокой частоты воспринимаются короткими волокнами основной мембраны, расположенными ближе к основанию улитки. При действии сложного звука возникают колебания различных волокон мембраны.
В современной интерпретации резонансный механизм лежит в основе теории места, в соответствии с которой в состояние колебания вступает вся мембрана. Однако максимальное отклонение основной мембраны улитки происходит только в определенном месте. При увеличении частоты звуковых колебаний максимальное отклонение основной мембраны смещается к основанию улитки, где располагаются более короткие волокна основной мембраны, – у коротких волокон возможна более высокая частота колебаний. Возбуждение волосковых клеток именно этого участка мембраны при посредстве медиатора передается на волокна слухового нерва в виде определенного числа импульсов, частота следования которых ниже частоты звуковых волн (лабильность нервных волокон не превышает 800 – 1000 Гц). Частота воспринимаемых звуковых волн достигает 20 000 Гц. Таким способом осуществляется пространственный тип кодирования высоты и частоты звуковых сигналов.
При действии тонов примерно до 800 Гц кроме пространственного кодирования происходит еще и временное (частотное) кодирование, при котором информация передается также по определенным волокнам слухового нерва, но в виде импульсов (залпов), частота следования которых повторяет частоту звуковых колебаний. Отдельные нейроны на разных уровнях слуховой сенсорной системы настроены на определенную частоту звука, т.е. каждый нейрон имеет свой специфический частотный порог, свою определенную частоту звука, на которую реакция нейрона максимальна. Таким образом, каждый нейрон из всей совокупности звуков воспринимает лишь определенные достаточно узкие участки частотного диапазона, не совпадающие между собой, а совокупности нейронов воспринимают весь частотный диапазон слышимых звуков, что и обеспечивает полноценное слуховое восприятие.
Правомерность этого положения подтверждается результатами протезирования слуха человека, когда электроды вживлялись в слуховой нерв, а его волокна раздражались электрическими импульсами разных частот, которые соответствовали звукосочетаниям определенных слов и фраз, обеспечивая смысловое восприятие речи.
Анализ интенсивности звукатакже осуществляется в слуховой сенсорной системе. При этом сила звука кодируется как частотой импульсов, так и числом возбужденных рецепторов и соответствующих нейронов. В частности, наружные и внутренние волосковые рецепторные клетки имеют разные пороги возбуждения. Внутренние клетки возбуждаются при большей силе звука, чем наружные. Кроме того, у внутренних клеток пороги возбуждения также различны. В связи с этим в зависимости от интенсивности звука меняются соотношение возбужденных рецепторных клеток кортиева органа и характер импульсации, поступающей в ЦНС. Нейроны слуховой сенсорной системы имеют различные пороги реакций. При слабом звуковом сигнале в реакцию вовлекается лишь небольшое число более возбудимых нейронов, а при усилении звука возбуждаются нейроны с меньшей возбудимостью.
Необходимо отметить, что кроме воздушной проводимости имеется костная проводимость звука,т.е. проведение звука непосредственно через кости черепа. При этом звуковые колебания вызывают вибрацию костей черепа и лабиринта, что приводит к повышению давления перилимфы в вестибулярном канале больше, чем в барабанном, так как перепонка, закрывающая круглое окно, эластична, а овальное окно закрыто стремечком. В результате этого происходит смещение основной мембраны, так же как и при воздушной передаче звуковых колебаний.
Определение локализации источника звукавозможно с помощью бинаурального слуха, т. е. способности слышать одновременно двумя ушами. Благодаря бинауральному слуху человек способен более точно локализовать источник звука, чем при монауральном слухе, и определять направление звука. Для высоких звуков определение их источника обусловлено разницей силы звука, поступающего к обоим ушам, вследствие различной их удаленности от источника звука. Для низких звуков важной является разность во времени между приходом одинаковых фаз звуковой волны к обоим ушам.
Определение местоположения звучащего объекта осуществляется либо путем восприятия звуков непосредственно от звучащего объекта – первичная локализация, либо путем восприятия отраженных от объекта звуковых волн – вторичная локализация, или эхолокация. При помощи эхолокации ориентируются в пространстве некоторые животные (дельфины, летучие мыши).
Слуховая адаптация– это изменение слуховой чувствительности в процессе действия звука. Она складывается из соответствующих изменений функционального состояния всех отделов слухового анализатора. Ухо, адаптированное к тишине, обладает более высокой чувствительностью к звуковым раздражениям (слуховая сенситизация). При длительном слушании слуховая чувствительность снижается. Большую роль в слуховой адаптации играет ретикулярная формация, которая не только изменяет активность проводникового и коркового отделов слухового анализатора, но и за счет центробежных влияний регулирует чувствительность слуховых рецепторов, определяя уровень их «настройки» на восприятие слуховых раздражителей.
Дата добавления: 2015-10-05; просмотров: 1829;