Строение тканей овощей и плодов
Технологические свойства овощей и плодов определяются составом и содержанием в них пищевых веществ и особенностями строения их тканей.
Ткань овощей состоит из тонкостенных клеток, такую ткань называют паренхимной. Содержимое клеток представляет собой полужидкую массу – цитоплазму, в которую погружены органеллы клетки – вакуоли, ядра, пластиды.
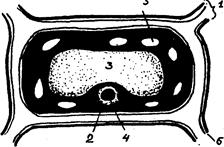
Рис. 1. Строение растительной клетки:
1 – клеточная стенка; 2 – цитоплазма;
3 – вакуоль; 4 – ядро; 5 – хлоропласты;
6 – клеточная оболочка.
Все органеллы клетки отделены от цитоплазмы мембранами, вакуоли окружены простой мембраной – тонопластом. Поверхность ядер, пластид и других цитоплазматических структур покрыта двойной мембраной. Цитоплазма на границе с клеточной оболочкой покрыта плазмолеммой (простая мембрана), которую можно увидеть при помещении клетки в солевой раствор. Вследствие разницы давления внутри клетки и вне ее происходит переход воды из клетки в окружающую среду, вызывающий плазмолиз – отделение цитоплазмы от клеточной оболочки.
Каждая растительная клетка покрыта оболочкой, представляющей собой первичную клеточную стенку. В отличие от мембран она характеризуется полной проницаемостью. Оболочки каждых соседних клеток скреплены при помощи срединных пластинок, образуя остов паренхимной ткани.
Поверхность отдельных экземпляров овощей и плодов покрыта покровной тканью – эпидермисом (плоды, наземные овощи) или перидермой (картофель, свекла, репа). Свежие отличаются повышенным содержанием воды, поэтому все структурные элементы их паренхимной ткани гидратированы. Способность овощей и плодов при достаточно высоком содержании влаги сохранять форму объясняется присутствием белков и углеводов, способных удерживать значительное количество влаги. Это обеспечивает достаточно высокое тургорное давление в тканях. Тургор – это состояние напряжения, возникающее вследствие давления содержимого клеток на их эластичные оболочки и давления оболочек на содержимое клеток.
2. Изменения углеводов клеточных стенок при тепловой кулинарной обработке: клетчатки, полуклетчатки, пектиновых веществ
Из углеводов в овощах содержатся моносахариды (глюкоза и фруктоза), дисахариды (сахароза, мальтоза) и полисахариды (крахмал, клетчатка, гемицеллюлозы, пектиновые вещества). Клеточные стенки составляют примерно 0,7-5% сырой массы овощей и плодов. В состав клеточных стенок и срединных пластинок входят в основном полисахариды (80-95%) – клетчатка, гемицеллюлозы и протопектин, поэтому их называют «углеводами клеточных стенок». Кроме углеводов в клеточных стенках содержатся азотистые вещества, лигнин, липиды, воска, минеральные вещества. Из азотистых веществ в клеточных стенках обнаружен белок, который в некоторых отношениях напоминает белок коллаген, выполняющий аналогичные функции в животных тканях, называют его экстенсином.
В начальный период тепловой кулинарной обработки овощей и плодов активизируются содержащиеся в них ферменты, вызывающие те или иные изменения пищевых веществ, затем ферменты инактивируются, цитоплазма и мембраны вследствие денатурации белков разрушаются и отдельные компоненты клеточного сока и других структурных элементов клетки получают возможность взаимодействовать друг с другом.
При тепловой обработке происходит размягчение овощей и плодов, обусловленное частичной деструкцией клеточных стенок, однако клеточные стенки не разрушаются даже при протирании овощей, так как обладают достаточной прочностью и эластичностью. При протирании ткань разрушается по срединным пластинкам, которые подвергаются большей деструкции. Глубоким изменения подвергаются углеводы клеточных стенок: гемицеллюлозы и пектиновые вещества, а также экстенсин, целлюлоза при тепловой обработке набухает.
Деструкция протопектина и гемицеллюлоз. Протопектин — полимер пектина — имеет сложную разветвленную структуру. Главные цепи его молекул состоят из остатков галактуроновых и полигалактуроновых кислот и сахара – рамнозы. Цепи галактуроновых кислот соединены друг с другом с помощью различных связей (водородных, эфирных, ангидридных, солевых мостиков), среди которых преобладают солевые мостики из двухвалентных ионов кальция и магния. Деструкции протопектина обусловлена, прежде всего, распадом водородных связей между этерифицированными остатками галактуроновой кислоты и хелатных связей с участием ионов Са++ и Мg++ между неэтерифицированными остатками полигалактуроновой кислоты. Происходит ионообменная реакция: ионы кальция и магния заменяются одновалентными ионами натрия и калия.
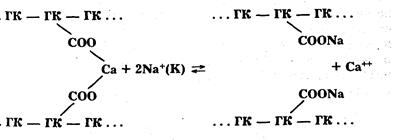
При этом связь между отдельными цепями галактуроновых кислот разрушается. Протопектин распадается, образуется растворимый в воде пектин, и овощная ткань размягчается. Реакция эта обратима. Чтобы она проходила в правую сторону, необходимо удалять ионы кальция из сферы реакции. В растительных продуктах содержатся фитин и ряд других веществ, связывающих кальций. Однако связывание ионов кальция (магния) не происходит в кислой среде, поэтому размягчение овощей замедляется. В жесткой воде, содержащей ионы кальция и магния, этот процесс также будет проходить медленно.
Таким образом, особенность механизма деструкции клеточных стенок различных овощей определяется, прежде всего, степенью этерификации остатков галактуроновой кислоты в протопектине. Степень этерификации полигалактуроновой кислоты в свекле 72%, моркови – 59%, капусте – 65%, у картофеля и кабачков – 40%.
В разных овощах скорость распада протопектина не одинакова. Поэтому варить можно все овощи, а жарить только те, у которых протопектин успевает превратиться в пектин, пока еще не вся влага испарилась (картофель, кабачки, помидоры, тыкву). Можно предположить, что жарке пригодны те овощи, у которых степень этерификации полигалактуроновых кислот близка к 40%. У моркови, репы, брюквы и некоторых других протопектин настолько устойчив, что они начинают подгорать раньше, чем достигнут кулинарной готовности.
При тепловой кулинарной обработке овощей наряду и параллельно с деструкцией протопектина протекает деструкция гемицеллюлоз также с образованием растворимых продуктов. Однако деструкция гемицеллюлоз начинается при более высоких температурах, чем деструкция протопектина – от 70 до 80ºС.
Деструкция структурного белка клеточных стенок – экстенсина. Экстенсин при тепловой обработке подвергается деструкции с образованием растворимых продуктов. Разрушение экстенсина начинается уже при температуре 50ºС, при этом несколько уменьшается и механическая прочность овощей.
Установлено, что свекла при варке до полуготовности и последующем охлаждении размягчается, то есть когда протопектин, гемицеллюлозы и пектин уже подверглись необходимой деструкции, но продукты деструкции еще удерживаются в связанном состоянии и не перешли в раствор, так как необходимая для этого влага в клеточных стенках отсутствует. При остывании корнеплодов набухшая целлюлоза и гемицеллюлозы частично восстанавливают структуру и выделяют поглощенную при набухании влагу, в результате такого обводнения клеточных стенок и дополнительного поступления влаги из клеток растворимые продукты деструкции протопектина, гемицеллюлоз и экстенсина переходят в раствор и прочность клеточных стенок понижается.
При охлаждении сваренных овощей и плодов эластичность клеточных оболочек понижается, а хрупкость возрастает. Это обусловливает неприятно вязкую консистенцию при протирании картофеля в охлажденном состоянии, при котором происходит разрушение клеточных стенок и клейстеризованный крахмал вытекает.
Дата добавления: 2015-11-18; просмотров: 4409;