Развитие нервной системы в онтогенезе.
Пренатальный период онтогенеза начинается с момента слияния мужских и женских половых клеток и образования зиготы. Зигота последовательно делится, образуя шаровидную бластулу. На стадии бластулы идет дальнейшее дробление и образование первичной полости — бластоцеля.
Затем начинается процесс гаструляции, в результате которого происходит перемещение клеток различными способами в бластоцель, с образованием двухслойного зародыша. Наружный слой клеток называется эктодерма, внутренний — энтодерма.
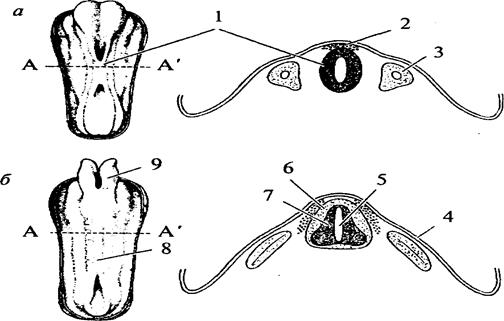
Рис. 12. Закладка нервной трубки (схематичное изображение и вид на поперечном срезе):А—А'— уровень поперечного среза; а — начальный этап погружения медуллярной пластинки и формирования нервной трубки: 1 — нервная трубка; 2 — ганглиозная пластина; 3 — сомит; б — завершение образования нервной трубки и погружение ее внутрь зародыша: 4 — эктодерма; 5 — центральный канал; 6 — белое вещество спинного мозга; 7 — серое вещество спинного мозга; 8 — закладка спинного мозга; 9 — закладка головного мозга
Внутри образуется полость первичной кишки — гастроцель. Это стадия гаструлы.
На стадии нейрулы образуются нервная трубка, хорда, сомиты и другие эмбриональные зачатки.
Зачаток нервной системы начинает развиваться еще в конце стадии гаструлы. Клеточный материал эктодермы, расположенный на дорсальной поверхности зародыша, утолщается, образуя пластинку которая ограничивается с боков валиками. Дробление клеток пластинки и валиков приводит к изгибанию пластинки в желоб, а затем к смыканию его краев и образованию нервной трубки.
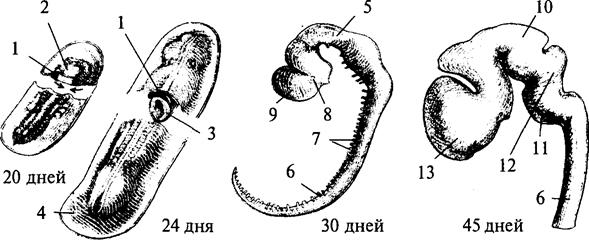
Рис. 13. Пренатальное развитие нервной системы человека:
1 — нервный гребень; 2 — нервная пластина; 3 — нервная трубка; 4 — эктодерма; 5 — средний мозг; 6 — спинной мозг; 7 — спинномозговые нервы; 8 — глазной пузырек; 9 — передний мозг; 10 — промежуточный мозг; 11 — мост; 12 — мозжечок; 13 — конечный мозг
Одновременно происходит погружение нервной трубки внутрь зародыша. Однородные первичные клетки стенки трубки дифференцируются на первичные нервные клетки (нейробласты) и исходные клетки нейроглии.
Клетки внутреннего, прилежащего к полости трубки, слоя выстилают просвет полостей мозга.
Все первичные клетки активно делятся, увеличивая толщину стенки мозговой трубки и уменьшая просвет нервного канала.
Нейробласты дифференцируются на нейроны. При дифференцировке нейробластов отростки удлиняются и превращаются в дендриты и аксон, которые на данном этапе лишены миелиновых оболочек.
Миелинизация начинается с пятого месяца пренатального развития и полностью завершается лишь в возрасте 5—7 лет.
На пятом же месяце появляются синапсы.
Миелиновая оболочка формируется в пределах ЦНС олигодендроцитами, а в периферической нервной системе — Шванновскими клетками.
В первые месяцы постнатального онтогенеза продолжается интенсивный рост аксонов и дендритов и резко возрастает количество синапсов в связи с развитием нейронных сетей.
Эмбриогенез головного мозга начинается с развития в передней части мозговой трубки двух первичных мозговых пузырей, возникающих в результате неравномерного роста стенок нервной трубки.
Затем в начале четвертой недели у зародыша второй пузырь делится на средний и ромбовидный пузыри.
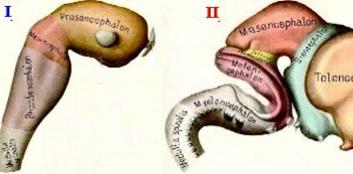
| Схема - головной мозг эмбриона: I - стадия трёх мозговых пузырей; II -стадия образования пяти отделов мозга (II). |
В нижней части переднего мозга выпячиваются обонятельные лопасти (из них развиваются обонятельный эпителий носовой полости, обонятельные луковицы и тракты). Из дорсолатеральных стенок переднего мозгового пузыря выступают два глазных пузыря. В дальнейшем из них развиваются сетчатка глаз, зрительные нервы и тракты.
На шестой неделе эмбрионального развития передний и ромбовидный пузыри делятся каждый на два и наступает пятипузырная стадия.
Передний пузырь — конечный мозг — разделяется продольной щелью на два полушария. Полость также делится, образуя боковые желудочки. Мозговое вещество увеличивается неравномерно, и на поверхности полушарий образуются многочисленные складки — извилины, отделенные друг от друга более или менее глубокими бороздами и щелями.
Каждое полушарие разделяется на четыре доли. Из мезенхимы, окружающей мозг зародыша, развиваются оболочки мозга. Серое вещество располагается и на периферии, образуя кору больших полушарий, и в основании полушарий, образуя подкорковые ядра.
Задняя часть переднего пузыря остается неразделенной и называется теперь промежуточным мозгом. Функционально и морфологически он связан с органом зрения. На стадии, когда границы с конечным мозгом слабо выражены, из базальной части боковых стенок образуются парные выросты — глазные пузыри, которые соединяются с местом их происхождения при помощи глазных стебельков, впоследствии превращающихся в зрительные нервы.
Наибольшей толщины достигают боковые стенки промежуточного мозга, которые преобразуются в зрительные бугры, или таламус. В вентральной области (гипоталамус) образуется непарное выпячивание — воронка, из нижнего конца которой происходит задняя мозговая доля гипофиза — нейрогипофиз.
Третий мозговой пузырь превращается в средний мозг, который развивается наиболее просто и отстает в росте. Стенки его утолщаются равномерно, а полость превращается в узкий канал — Сильвиев водопровод, соединяющий III и IV желудочки. Из дорсальной стенки развивается четверохолмие, а из вентральной — ножки среднего мозга.
Ромбовидный мозг делится на задний и добавочный. Из заднего формируется мозжечок — сначала червь мозжечка, а затем полушария, а также мост. Добавочный мозг превращается в продолговатый мозг. Стенки ромбовидного мозга утолщаются — как с боков, так и на дне, только крыша остается в виде тончайшей пластинки.
В результате неравномерного развития мозговых пузырей мозговая трубка начинает изгибаться.
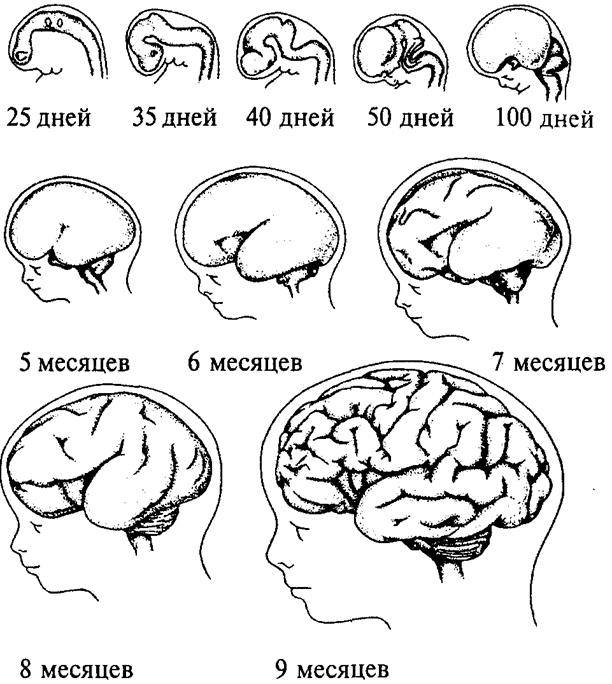
Рис. 14. развитие головного мозга .
Постнатальный онтогенез нервной системы человека начинается с момента рождения ребенка. Головной мозг новорожденного весит 300 - 400 г. Вскоре после рождения прекращается образование из нейробластов новых нейронов, сами нейроны не делятся. Однако к восьмому месяцу после рождения вес мозга удваивается, а к 4 - 5 годам утраивается. Масса мозга растет в основном за счет увеличения количества отростков и их миелинизации. Максимального веса мозг мужчин достигает к 20 - 29 годам, а женщин к 15 - 19. После 50 лет мозг уплощается, вес его падает и в старости может уменьшиться на 100 г.
У новорожденного вес спинного мозга 10 г, а отношение спинного мозга к головному 1:100 (у взрослых 1:49). В первый год жизни спинной мозг растет быстро, а дифференцировка его нейронов слабо выражена. Проводящие пути и спинномозговые нервы не покрыты миелиновой оболочкой. Отсюда широкая иррадиация возбуждения по сегментам спинного мозга.
На втором этапе развития ребенка рост спинного мозга отстает от позвоночника. В результате этого сегменты спинного мозга, в нижнем его отделе, перестают соответствовать по проекции позвонкам. Вес его достигает 14 г. На этом этапе заканчивается миелинизация всех спинномозговых нервов и восходящих (афферентных) проводящих путей. Эфферентные проводящие пути еще не полностью покрыты миелиновой оболочкой. Следствием этого является неточность координации спинномозговых рефлексов. Так, например, в два года нет еще дифференцировки ходьбы от бега. Ребенок быстро перебирает ногами, шаги у него короткие. Только к концу этапа наступает дифференцировка ходьбы от бега и улучшается координация спинномозговых рефлексов.
Полностью заканчивается развитие спинного мозга на 3 этапе развития.
Продолговатый и задний мозг растут быстро. Все основные центры заднего мозга сформированы на первом году жизни. Однако их функция еще не совершенна, так как их регуляция осуществляется в основном за счет безусловных рефлексов, т.е. по принципу отклонения.
К концу 3 года его нейроны приобретают форму взрослого человека и отличаются только размерами (тела нейронов еще меньше, а аксоны и дендриты короче, чем у взрослых), поэтому их функции почти как у взрослых.
Мозжечоку новорожденного весит 20 г, а к концу года – 90 г. Червь растет быстрее полушарий. В коре полушарий как и у взрослых 3 слоя клеток, но они меньше чем у взрослых. Дифференцировка слоев начинается с первых месяцев после рождения, но к концу года еще остается много не дифференцированных нейронов. Поэтому функции мозжечка не совершенны, движения не координированы, так как нет точного согласования скорости сокращения различных мышц.
Начиная со второго этапа развития мозжечок растет равномерно, его клетки заканчивают дифференцировку и заканчивают свое развитие на 3 этапе развития.
Средний мозг по отношению к взрослым у новорожденного составляет 40%, к третьему году – 50%. Нейроны четверохолмия не дифференцированы. Функции среднего мозга не совершенны, движения не точны из-за несогласованности двигательного и тонического компонентов. Поздно-тонические, выпрямительные и стато-кинетические рефлексы не координированы. У новорожденного глазные яблоки могут даже двигаться одновременно в разные стороны. Сторожевые рефлексы (зрительные и слуховые ориентировочные рефлексы) слабо выражены и не точны. Отсутствует координация сокращений и перераспределения тонуса мышц пальцев рук.
На третьем этапе развития ребенка средний мозг составляет 90% от взрослого. Заканчивается дифференцировка почти всех его отделов. Однако, наблюдается несовершенство в координации движений пальцев рук и ребенку с большим трудом даются такие двигательные навыки как рисование, письмо, т.е. двигательные навыки требующие точной дифференцировки в движениях пальцев рук.
Ретикулярная формация ствола мозга занимает относительно меньший объем, чем у взрослых. Нейроны в ней расположены более тесно, их аксоны не покрыты миелиновой оболочкой. Многие нейроны еще не закончили своей дифференцировки. Активация больших полушарий слабая и их возбудимость понижена (новорожденный спит до 22 часов в сутки). Заканчивается дифференцировка нейронов ретикулярной формации на 3 этапе развития.
Промежуточный мозг растет относительно медленно. Так, размеры зрительных бугров составляют только 50% от взрослого. Нет еще дифференцировки переключающих и ассоциативных ядер. Нет миелинизации проводящих путей. Отсюда несовершенны функции промежуточного мозга. Нет четкости в передаче информации в кору больших полушарий, отсутствует контроль со стороны высших вегетативных центров, имеется несовершенство терморегуляции.
На втором этапе развития в зрительных буграх происходит дифференцировка нейронов на две группы ядер:
i. переключающие ядра;
ii. ассоциативные ядра.
Афферентные пути от переключающих ядер к сенсорным зонам коры больших полушарий полностью покрыты миелиновой оболочкой, а афферентные пути от ассоциативных ядер к ассоциативным зонам больших полушарий еще не полностью покрыты миелиновой оболочкой. Отсюда несовершенен еще корковый анализ и синтез, особенно комплексных раздражителей. Неспецифические ядра зрительных бугров еще недифференцированы. Отсюда у детей этого этапа развития активное внимание практически отсутствует.
В гипоталамусе внутри ядер нейроны не полностью дифференцированы, их аксоны не покрыты миелиновой оболочкой, поэтому сохраняется несовершенство контроля над вегетативной нервной системой.
Завершается формирование промежуточного мозга к концу 4 этапа развития.
Подкорковые ядра составляют 20% по отношению к взрослым. Дифференцировка нейронов слабая, миелинизация проводящих путей неполная. Несовершенство функций проявляется в неловкости двигательных актов. К третьему этапу развития подкорковые ядра составляют 80% от взрослых и завершают свое развитие к концу 4 этапа.
Большие полушария. Формообразование больших полушарийна первом году жизни идет медленными темпами, и они по своей форме значительно отличаются от взрослого человека. Так, у детей первого года жизни слабо развит лобный отдел больших полушарий, борозды и извилины малы, мелки. Особенно плохо выражены борозды третьего порядка.
На II этапе поверхность больших полушарий становится похожей на таковую у взрослого человека.
Замедлена на первом этапе развитие и дифференцировка нейронов коры головного мозга. Хотя в коре больших полушарий уже у новорожденного имеется такое же количество нейронов (15 млрд.), как и у взрослого, но на протяжении почти всего 1 этапа развития форма нейронов остается несовершенной. Большинство нейронов имеют овальную форму со слабо выраженными дендритами и короткими аксонами. В связи с этим у новорожденного нет деления нейронов на слои и только к концу первого года жизни в коре больших полушарий начинают определяться 6 слоев нейронов, отличающихся друг от друга, как по форме, так и по размерам. Нейроны приобретают форму типичную для нейронов коры больших полушарий взрослого человека (пирамидную, звездчатую, треугольную, веретенообразную и т.д.). Однако корковые нейроны еще несколько меньших размеров, чем у взрослых, их дендриты еще коротки, а аксоны не покрыты миэлиновой оболочкой.
На III этапе заканчивается дифференцировка тел и дендритов корковых нейронов. Однако их аксоны еще не полностью покрыты миелиновой оболочкой. Хотя на этом этапе развития и устанавливаются связи между слоями коры больших полушарий, однако передача информации из одного слоя в другой не совершена, так как не все аксоны покрыты миелиновой оболочкой. Не покрыты миелиновой оболочкой и ассоциативные пути, соединяющие ассоциативные зоны больших полушарий, следовательно, не совершенен корковый анализ и синтез. Сенсорные и моторные зоны больших полушарий развиты почты также, как и у взрослых, а ассоциативные и моторные зоны на 80% по сравнению со взрослыми.
В связи с незрелостью коры больших полушарий, функция ее несовершенна, контроль над подкорковыми образованиями слабый. Так, новорожденный 22 часа в сутки спит, т.е. большую часть суток кора вообще не функционирует. Отсутствие корковых влияний на сегменты спинного мозга в первые месяцы жизни ребенка приводит к хаотичности движений (ребенка пеленают). Только со 2-3-го месяца жизни кора больших полушарий начинает более или менее оказывать постоянные регулирующие влияния на подкорковые образования. Однако эти влияния еще слабы, несовершенны.
На II этапе развития морфологическое и функциональное созреванием нейронов больших полушарии значительно совершенствуется. Так, благодаря корковому контролю над спинным мозгом более совершенными становятся произвольные движения (ребенок ходит, бегает, прыгает и т.д.). Достаточно хорошо развиты на этом этапе анализаторы, в том числе и дистантные (зрительный, слуховой и обонятельный). Быстро начинают образовываться временные связи во второй сигнальной системе и к концу 3-го года жизни ребенок знает уже около 1 000 слов.
Однако кора больших полушарий на II этапе развития ребенка все еще остается функционально недоразвитой, контроль ее над нижележащими отделами ЦНС несовершенен. Особенно это касается эмоциональной сферы ребенка. Ребенок "живет чувствами" (смеется на похоронах, плачет в гостях и т.д.).
Функциональная незрелость коры больших полушарий проявляется и в относительной слабости процессов торможения и в относительной легкости иррадиации процессов возбуждения (ребенок быстро перевозбуждается во время игры, плохо засыпает, спит беспокойно и т.д.). На III этапе развития нет еще контроля коры над эмоциями (можно узнать правду "по глазам").
Поскольку некоторые ученые функцию мышления связывают с лобными долями и, в частности, с 3-м слоем коры больших полушарий, а также с ассоциативными зонами больших полушарий, можно говорить о морфологической и, следовательно, функциональной незрелости больших полушарий на IV этапе развития ребенка, о несовершенстве коркового анализа и синтеза. Как известно, в этом возрасте мышление у детей все еще остается конкретным, хотя они уже и начинают пользоваться абстрактными понятиями. То, что взрослому кажется "само собой разумеющимся", "само собой вытекающим ...", для ребенка 7-10 лет может быть непонятным.
Отсюда преподавание должно строиться не только на словесном логическом доказательстве, но обязательно с использованием наглядностей (таблицы, рисунки, модели, опыты и т.д.), с обязательным знакомством ребенка с конкретными явлениями и предметами.
В связи с функциональной незрелостью корковых нейронов наблюдается и их относительная малая работоспособность. Наряду с этим наблюдается преобладание процессов возбуждения над процессами торможения, легкая и широкая иррадиация возбуждения. Концентрация же возбуждения и торможения затруднена. Так, например, продолжительность активного внимания (которое физиологически можно представить как концентрированный очаг возбуждения с индуцированным вокруг него торможением) в 7-8 лет равна 15-20 мин., а в 9-10 лет 20-25 мин.
Младшие школьники легко возбуждаются и перевозбуждаются во время перемен и долго не могут успокоиться в начале следующего урока. В связи с относительной слабостью процессов торможения, для младшего школьника неподвижно сидеть во время урока в течение 45 мин. - это большой нервный труд. Поэтому во время перемены детям необходимо давать разрядку, снимать торможение с двигательной зоны больших полушарий, т.е. дать возможность младшим школьникам побегам, поиграть в подвижные игры. Однако при этом надо следить, чтобы они не перевозбудились.
На IV этапе кора больших полушарий берет под контроль эмоции (уже нельзя узнать правду "по глазам"). Совершенствуется мышление, которое становиться словесным с использованием абстрактных понятий.
Только к 13 -16 годам заканчивается миэлинизация ассоциативных путей. Усиливаются процессы торможения, совершенствуется концентрация процессов возбуждения и торможения.
Так, например, продолжительность активного внимания в этом возрасте достигает 30-40 мин.
К 17 -18 годам морфологическое развитие больших полушарий оканчивается, но функциональное совершенствование продолжается и у взрослых людей. Так, большинство ученых считают, что функциональное совершенствование больших полушарий продолжается до 50-60 лет.
Дата добавления: 2015-08-14; просмотров: 3111;