Рецепторно-канальные комплексы типа I
Как показано на рисунке 16, в комплексе типа I рецепторное звено встроено в эффекторное звено – стенку белкового канала. Вместе они образуют рецептор-канал, в котором функцию сопрягающего звена выполняет непосредственный контакт, что обеспечивает быстроту физиологического ответа на действие лиганда.
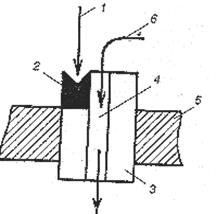
| Рис.16. Схема организации рецепторно-канального комплекса типа I. 1 – лиганд; 2 – рецепторное звено (участок) в канальном белке; 3 – эффектор (стенка канала); 4 – ионный канал; 5 – плазмолемма; 6 – ион. |
Известны четыре вида рецептор-каналов: никотиновый холинорецептор (Н-холинорецептор), гамма-аминомасляный, глутаматный и глициновый. В качестве примера рассмотрим два из них: Н-холиновый и глутаматный.
· Н-холинорецепторы
К одному и тому же нейромедиатору ацетилхолину существует два типа рецепторов, один из которых возбуждается никотином (Н-холинорецепторы), другой – мускарином (М-холинорецепторы).
Н-холинорецептор является составной частью быстродействующего рецептор-канала (Рис.17). М-холинорецепторы управляют своими эффектор-каналами посредством специализированных белков, что значительно снижает быстродействие.
Н-холинорецептор имеет грибовидную форму. Он пронизывает клеточную мембрану насквозь, выступая концами за наружную и внутреннюю ее стороны. Сверху молекула Н-холинорецептора имеет вид неправильной по форме розетки из пяти субъединиц, расположенных вокруг центрального электронно-плотного образования, ограничивающего ионный канал.
В Н-холиновом рецептор-канале выделяют два функционально различных компонента – узнающий центр и ионный канал. Узнающий центр – участки альфа-субъединиц, выступающие над поверхностью клеточной мембраны и обладающие способностью к опознанию и связыванию молекул ацетилхолина. Эти участки и составляют собственно Н-холинорецептор.
Молекулярный по своей организации рецептор встроен в стенку исполнительной части - канального белка, так что в целом все образование представляет собою рецепторно-эффекторный комплекс, предназначенный для генерирования потенциала действия в плазмолемме. Н-холинорецепторный комплекс является первым звеном Н-холинореактивной системы более высокого уровня, осуществляющей функцию сокращения поперечнополосатой мышцы.
| Рис.17. Схематическое изображение Н-холинового рецептор-канала (трехмерное изображение). I – грибовидный рецептор-канал; II – плазматическая мембрана. | 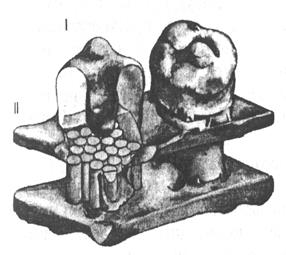
|
В фармакологии и нейротоксикологии известно большое число химиопрепаратов, которые специфически взаимодействуют с рецепторными участками, ионными каналами, а также с отдельными звеньями на пути дальнейшего прохождения сигналов к эффекторам, обеспечивающим конечные биохимические и физиологические ответы клетки. Такие вещества используются для изучения холинореактивных и других систем, а также в лечебных целях.
· Глутаматные рецепторы
Ряд аминокислот (глутаминовая, аспарагиновая, гамма-аминомасляная, тауриновая и др.), кроме энергетических, выполняют еще и медиаторные функции, оказывая на клетки-мишени возбуждающие и тормозные действия. Глутамат (анион глутаминовой кислоты) является одним из основных возбуждающих нейропередатчиков в нервной системе.
Наибольшее количество глутаматных рецепторов находится в коре больших полушарий, гиппокампе, полосатом теле, среднем мозге и в гипоталамусе.
Глутаматный рецептор-канал состоит из четырех белковых субъединиц, образующих эллипсоидной формы комплекс с центрально расположенным воронкообразным каналом для селективного пропуска ионов натрия, калия, хлора, кальция (Рис.18).
Вполне вероятно, что природа ионов, проходящих через канал, определяется типом нейронов и их функциональной специализацией. Рецепторный участок встроен в одну из субъединиц.
В отличие от Н-хлоинорецепторов, стабильно закрепленных в клеточной мембране, белковые субъединицы глутаматных рецептор-каналов в отсутствие лигандов находятся в диссоциированном состоянии и свободно плавают внутри мембраны или на ее поверхности.
При поступлении глутамата субъединицы, по-видимому, ассоциируются, образуя рецептор-канал, причем непосредственно над предсуществующим трансмембранным ионным каналом, выступая над ним в виде пристройки и стимулируя его раскрытие. Следует подчеркнуть, что строгих доказательств в пользу вышеупомянутых событий в литературе еще нет. Появляются сведения о том, что имеются разные типы глутаматных рецепторов: быстродействующие (тип I) и медленнодействующие (тип II).
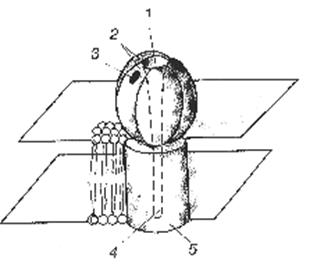
| Рис.18. Глутаматные рецепторы. 1 – воронкообразный канал рецептор-канале; 2 – белковые субъединицы; 3 – участокопознанияи связывания глутамата на одной из субъединиц; 4 –канал внутри трансмембранного белка (5). |
Описанный выше рецептор-канал является быстродействующим. Медленнодействующие глутаматные рецепторы участвуют в реализации химического сигнала через систему внутриклеточных посредников: циклических нуклеотидов и ионов кальция, однако соотношение их с быстродействующими и преимущественные места расположения еще не ясны.
Механизм действия глутамата в системе головного мозга заключается в индукции состояния возбуждения нейронов и их ансамблей. Связывание глутамата с рецептор-каналом вызывает в постсинаптической мембране волну возбуждения, которая распространяется по всему нейрону и по синапсам перекидывается на соседние нейроны.
Глутаматные рецепторы широко представлены в структурах головного мозга, ответственных за проявление высших психических и двигательных функций. Ряд нервно-психических заболеваний, таких, как хорея Гентингтона, шизофрения, эпилепсия, связаны с количественными и качественными изменениями глутаматергических систем.
Дата добавления: 2015-08-08; просмотров: 1537;