Структуры лимбической системы
К миндалине и гиппокампу идут пути от височной доли коры, передающие информацию от зрительной, слуховой и соматической сенсорной систем. Установлены связи ЛС с лобными долями коры переднего мозга. Наконец, в пределах ЛС идентифицированы сложные циклические связи, создающие условия для циркуляции возбуждения по сложным круговым путям. Примером такой циклической связи может служить так называемый круг Папеса, идущий от гиппокампа через свод – мамиллярное тело – переднее ядро таламуса – кору поясной извилины и обратно к гиппокампу.
Очевидно, сложность связей и внутренней организации ЛС свидетельствует об ее участии в интеграции функций новой коры и стволовых образований головного мозга.
Функции ЛС:
1. Ряд соматических и вегетативных функций, например торможение дыхания, повышение кровяного давления, шевеление губами и т.д.;
2. Формирование эмоциональных и поведенческих реакций;
3. Участие в механизмах памяти.
В 1937 г. американский невропатолог Д.В. Папес выдвинул гипотезу, согласно которой структуры мозга, расположенные в древней и старой коре, образуют единую систему (круг Папеса), ответственную за осуществление врожденных поведенческих актов и формирование эмоций. В 1952 г. другой американский исследователь, П.Д. Мак-Лин, развивая идеи Папеса, ввел понятие лимбической системы. Этим термином исследователь обозначил сложную функциональную систему, обеспечивающую постоянство внутренней среды и контроль видоспецифических реакций, направленных на сохранение вида. Теоретические и практические разработки Папеса и Мак-Лина послужили мощным толчком для дальнейших исследований в этой области.
Эти исследования показали, что локальное раздражение различных отделов ЛС вызывает разнообразные вегетативные эффекты и влияет на деятельность внутренних органов. Так, раздражение миндалины приводит к изменениям частоты сердечных сокращений, дыхательных движений, сосудистого тонуса. В ряде случаев раздражение миндалин влияет на деятельность пищеварительного тракта, изменяя перистальтику тонкого кишечника, стимулируя секрецию слюны, произвольное жевание и глотание. Описано влияние миндалин на сокращение мочевого пузыря и пилоэрекцию. Все эти разнообразные реакции могут иметь различный знак и характеризоваться активацией или угнетением вегетативных функций.
Вместе с тем высказывается предположение о том, что регулирующие влияния миндалины опосредованы нижележащими вегетативными центрами, в частности, гипоталамусом.
Скорее всего, изменяя в ту или иную сторону возбудимость гипоталамических центров, ЛС определяет знак соответствующей вегетативной реакции. Так формируется многоэтажная, построенная по иерархическому принципу система управления вегетативной сферой, интегрирующая вегетативные и соматические реакции.
Как известно, эмоциональная окраска поведенческих реакций определяется не только вегетативными компонентами, но и соответствующими эндокринными сдвигами. В этом плане представляет интерес данные о влиянии ЛС на деятельность желез внутренней секреции. Установлено, что длительное (60 мин) раздражение ядер миндалины у обезьян вызывает повышение содержания кортикостероидов в плазме крови. Низкочастотное (12-36 имп/с) раздражение гиппокампа, напротив, уменьшает содержание кортикостероидов, которые выделяются надпочечниками при действии стресовых раздражителей. Очевидно, что нисходящие влияния этих структур ЛС на гипоталамус, а через него на гипофиз изменяют продукцию АКТГ (адренокортикотропного гормона), который регулирует активность надпочечников.
Таким образом, изменяя гормональный фон, ЛС в естественных условиях может участвовать в формировании побуждений к действию (мотиваций) и регулировать реализацию самих действий, направленных на устранение побуждения, усиливая или ослабляя эмоциональные факторы поведения.
Сведения об участии ЛС в формировании эмоций неоднозначны. Наибольшее их количество относится к роли миндалины и поясной извилины в этих процесах. При локальном электрическом раздражении миндалины могут быть получены эмоциональные реакции типа страха, гнева, ярости и агрессии. Двустороннее удаление височных долей вместе с миндалиной и гиппокампом вызывает у обезьян целый ряд сдвигов в эмоциональной сфере. Как правило, агрессивные обезьяны после этой операции становятся спокойными и доверчивыми. У животных наблюдается гиперорализм, когда все незнакомые предметы без разбора запихиваются в рот.
Удаление височных долей вызывает у обезьян гиперсексуальность, причем их половая активность может быть направлена даже на неодушевленные предметы. Послеоперационный синдром также сопровождается так называемой психической слепотой. Животные утрачивают способность правильной оценки зрительной и слуховой информации. Так, обезьяны без разбора исследуют все, даже опасные для себя предметы. Возникновение психической слепоты связывают с послеоперационным нарушением передачи сенсорной информации от височной доли к гипоталамусу.
Согласно теории Папеса, сенсорные пути на уровне таламуса расходятся, причем один путь идет в проекционные зоны коры, где обеспечивается восприятие, а второй – в лимбическую систему. Вероятно, в этой системе происходит оценка поступающей информации, ее сопоставление с субъективным опытом и запуск соответствующих эмоциональных реакций через гипоталамические структуры. Информация об аффективном состоянии организма может поступать от стволовых структур в новую кору либо прямо от миндалины, либо через поясную извилину, которая связана с лобными, теменными и височными долями коры полушарий большого мозга. Вероятно, эти пути связаны с восприятием эмоционального фона, однако детальные механизмы этого восприятия еще нуждаются в разработке.
Как уже отмечалось выше, ЛС принимает участие в запуске тех эмоциональных реакций, которые уже апробированы в ходе жизненного опыта. В этом плане привлекают внимания исследования, констатирующие участие ЛС в процессах сохранения памяти. Так, например, удаление гиппокампов вызывает у людей полное выпадение памяти на недавние события. Электрическое раздражение гиппокампальной извилины во время хирургических операций может сопровождаться у больного появлением мимолетных воспоминаний. Двустороннее удаление гиппокампа у обезьян и крыс приводит к нарушению способности усваивать новые навыки. Описанные факты привели к заключению, что гиппокамп играет определенную роль в процессах памяти и послужили толчком для дальнейших исследований этой структуры.
Важно, что большинство исследователей склоняется к тому, что функции памяти не является весьма прерогативой какой-либо одной структуры, а обусловлены содружественными действиями многих центров головного мозга. Существенным звеном в этой системе являются связи гиппокампа с новой корой.
Функциональная роль этих связей подтверждается физиологическими экспериментами. При одновременной регистрации электрической активности гиппокампа и новой коры наблюдаются реципрокные взаимоотношения между ними. Когда в гиппокампе возникает медленноволновый тета-ритм, в новой коре доминирует высокочастотная низкоамплитудная активность и, наоборот, медленноволновой активности энцефалограммы соответствует высокочастотная активность гиппокампа.
Наиболее выраженное усиление тета-ритма в гиппокампе обнаруживается на начальных стадиях выработки условного рефлекса и сочетается с состоянием настороженности и сосредоточения внимания при формировании ориентировочной реакции.
Удаление гиппокампа у животных нарушает процессы внутреннего торможения и снижает способность к погашению потерявших свое адаптивное значение условно-рефлекторных реакций. Одновременно затрудняется упрочнение условного рефлекса в связи с резким усилением ориентировочной реакции. Следовательно, гиппокамп, как, впрочем, и другие структуры ЛС, существенно влияет на функции неокортекса и на процессы научения. Это влияние осуществляется в первую очередь за счет создания эмоционального фона, который в значительной степени отражается на скорости образования любого условного рефлекса.
Таким образом, ЛС является важным отделом головного мозга, функционально связанным с новой корой и стволовыми структурами, образующими вместе систему координации висцеральных и соматических функций организма.
Мантия полушарий
Мантия полушарий внутри состоит из белого вещества, снаружи покрытого серым – корой. Вся поверхность коры «изрезана» бороздами, между которыми располагаются извилины.
Оба полушария разделяются на 4 доли - лобную, теменную, височную и затылочную – тремя основными бороздами:
1. Латеральная борозда (Сильвиева) является передневерхней границей височной доли;
2. Центральная борозда разделяет полушария на передний и задний отделы, отграничивая лобную долю от теменной;
3. Теменно-затылочная борозда отделяет теменную долю от затылочной.
Общая поверхность площади коры полушарий составляет 1800 см2, причем 2/3 поверхности находится в глубине борозд, а 1/3 - на видимой поверхности.
На наружной поверхности лобной доли различают четыре извилины: вертикальную (прецентральную) и три горизонтальные (верхнюю, среднюю и нижнюю). Вертикальная извилина заключена между центральной и прецентральной бороздами. Верхняя лобная извилина расположена выше верхней лобной борозды, средняя - между верхней и нижней лобными бороздами, а нижняя - между нижней лобной и сильвиевой. На нижней (базальной) поверхности лобных долей различают прямую и орбитальную извилины, которые образованы обонятельной и орбитальными бороздами. Прямая извилина расположена между внутренним краем полушария и обонятельной бороздой. В глубине обонятельной борозды лежат обонятельная луковица и обонятельный тракт. В извилинах лобной доли сконцентрировано несколько функционально важных центров. Функция лобных долей связана с организацией произвольных движений, двигательных механизмов речи, регуляцией сложных форм поведения, процессов мышления.
Теменная доля отделена от лобной доли центральной бороздой, от височной - сильвиевой бороздой, от затылочной - воображаемой линией от верхнего края теменно-затылочной борозды до нижнего края полушария.
В теменной доле на наружной поверхности различают вертикальную постцентральную извилину и две горизонтальные дольки-верхнетеменную и нижнетеменную. Постцентральная извилина ограничена центральной и постцентральной бороздами, верхняя теменная долька расположена кверху от горизонтальной внутритеменной борозды, а нижняя - книзу от внутритеменной борозды. Часть нижней теменной дольки, расположенную над задним отделом сильвиевой борозды, называют надкраевой извилиной, а часть, окружающую восходящий отросток верхней височной бороздой, - угловой извилиной.
Функция теменной доли в основном связана с восприятием и анализом чувствительных раздражений, пространственной ориентации, регуляцией целенаправленных движений.
Височная доля отделена от лобной и теменной долей сильвиевой бороздой. На наружной поверхности височной доли различают верхнюю, среднюю и нижнюю извилины. Верхняя височная извилина располагается между сильвиевой и верхней височной бороздами, средняя - между верхней и нижней височными бороздами, нижняя - книзу от нижней височной - борозды. На нижней (базальной поверхности) височной доли находится латеральная затылочно-височная извилина, граничащая с нижней височной извилиной, а более медиально - извилина гиппокампа.
Функция височной доли связана с восприятием слуховых, вкусовых, обонятельных ощущений, анализом и синтезом речевых звуков, механизмами памяти.
Затылочная доля занимает задние отделы полушарий. На наружной поверхности затылочная доля не имеет четких границ, отделяющих ее от теменной и височной долей. На внутренней поверхности полушария теменную долю от затылочной ограничивает теменно-затылочная борозда. Внутреннюю поверхность затылочной доли разделяет шпорная борозда на клин (долька треугольной формы) и язычную извилину. Функция затылочной доли связана с восприятием и переработкой зрительной информации.
На внутренней поверхности полушарий над мозолистым телом находится поясная извилина, которая перешейком позади мозолистого тела переходит в парагиппокампову извилину. Поясная извилина вместе с парагиппокамповой извилиной составляет сводчатую извилину. На внутренней поверхности полушарий расположены участки коры, входящие в комплекс образований, относящихся к двум тесно связанным между собой функциональным системам-обонятельному мозгу и лимбической системе.
Строение мозговой коры
Кора в основном состоит из 6 слоев:
1. Молекулярный слой – содержит небольшое количество мелких ассоциативных клеток;
2. Наружный зернистый слой – в его состав входят мелкие нейроны;
3. Пирамидный слой – самый широкий из всех слоев коры – состоит из малых и средних пирамидных клеток;
4. Внутренний зернистый слой – образован мелкими звездчатыми нейронами;
5. Ганглиозный слой – образован крупными пирамидными нейронами, наибольшего размера достигающих в прецентральной извилине.
6. Слой полиморфных клеток – здесь находятся нейроны преимущественно веретенообразной формы. Этот слой граничит с белым веществом.
Учение о структурных особенностях строения коры головного мозга носит название архитектоники. Клетки коры больших полушарий менее специализированы, чем нейроны отдельных отделов мозга, тем не менее, определенные их группы анатомически и физиологически связаны с теми или иными специализированными отделами мозга. Из 6 слоев клеток коры верхние слои развиты у человека наиболее сильно. Нижние слои коры имеют связи с периферическими рецепторами (IV слой) и с мускулатурой (V слой) и носят название «первичных» или «проекционных» корковых зон, вследствие их непосредственной связи с периферическими отделами анализатора. Над «первичными» зонами надстраиваются системы «вторичных» зон (II и III слои), в которых преобладают ассоциативные связи с другими отделами коры, поэтому они называются проекционно-ассоциативными.
Мозг как субстрат психических процессов представляет собой единую систему, единое целое, состоящее, однако, из различных участков и зон, которые выполняют различную роль в реализации психических функций.
Морфологическая и функциональная неоднородности коры больших полушарий тесно связаны между собой. По данным Института мозга, описаны следующие цитоархитектонические поля коры больших полушарий: постцентральная область (поля 1, 2, 3, 43), прецентральная (поля 4, 6), лобная (поля 8-12, 44-47), теменная (поля 5, 7, 39, 40), височная (поля 20-22, 36-38, 41, 41), затылочная (17-19) и поясная область (поля 23-25, 31-33).
Все данные (анатомические, физиологические и клинические) свидетельствуют о ведущей роли коры больших полушарий в мозговой организации психических процессов.
В нейропсихологии на основе анализа нейропсихологических данных (т.е. изучения нарушений психических процессов при различных локальных поражениях мозга) была разработана общая структурно-функциональная модель мозга как субстрата психической деятельности. Эта модель, предложенная А. Лурия, характеризует наиболее общие закономерности работы мозга как единого целого и является основой для объяснения его интегративной деятельности. Согласно данной модели весь мозг может быть подразделен на три основных структурно-функциональных блока: а) энергетическии блок, или блок регуляции уровней активности мозга; б) блок приема, переработки и хранения экстероцептивной (исходящей извне) информации; в) блок программирования, регуляции и контроля над протеканием психической деятельности. Каждая высшая психическая функция осуществляется при участии всех трех блоков, каждый из которых вносит свой вклад в ее реализацию. Блоки характеризуются определенными особенностями строения, физиологическими принципами, лежащими в основе их работы, и той ролью, которую они играют в осуществлении психических функций.
Первый энергетический блок регулирует два типа процессов активации: общие генерализованные изменения активации мозга, являющиеся основой различных функциональных состояний, и локальные избирательные активационные изменения, необходимые для осуществления высших психических функций.
Функциональное значение первого блока в обеспечении психических функций, прежде всего, состоит, как уже говорилось выше, в регуляции процессов активации, â обеспечении того общего активационного уровня, на котором разыгрываются все психические функции, в поддержании общего тонуса ЦНС, необходимого любой психической деятельности. Этот аспект работы первого блока имеет непосредственное отношение к процессам внимания, а также сознания в целом. Помимо общих неспецифических активационных функций первый блок мозга непосредственно связан с процессами памяти, с запечатлением, хранением и переработкой разномодальной информации.
Второй блок - блок приема, переработки è хранения информации включает в себя основные анализаторные системы: зрительную, слуховую и кожно-кинестетическую, корковые отделы (зоны) которых расположены в задних отделах больших полушарий.
Все три анализаторные системы организованы по общему принципу: они состоят из периферического (рецепторного), проводникового и центральных отделов. Проводниковый отдел анализаторов обычно включает несколько уровней. Центральный отдел анализаторов - кора больших полушарий. В целом анализаторы - это аппараты, подготавливающие ответы организма на внешние раздражители.
Кора задних отделов больших полушарий обладает рядом общих черт, позволяющих объединить различные уровни анализаторных систем в единый блок мозга. Â коре задних отделов мозга выделяют первичные, вторичные и третичные поля, а по терминологии И.П. Павлова - «ядерные зоны анализаторов» и «периферию». В ядерную зону зрительного анализатора входят 17, 18 и 19-е поля, в ядерную зону кожно-кинестетического анализатора - 3, 1, 2-е, частично 5-е поле, в ядерную зону звукового анализатора - 41, 42 и 22-е поля, из них первичными полями являются - 17, 3 и 41-е. Остальные - вторичные.
Вторичные корковые поля осуществляют синтез раздражителей, функциональное объединение различных анализаторных зон, принимая непосредственное участие в обеспечении различных гностических видов психической деятельности.
Третичные поля коры больших полушарий находятся вне «ядерных зон» анализаторов. К ним относятся верхнетеменная область (7 и 40), нижнетеменная (39), средневисочная область (21 и частично 37). Функциональное значение третичных полей коры многообразно. С их участием осуществляются сложные надмодальностные виды психической деятельности - символической, речевой, интеллектуальной.
Третий структурно-функциональный блок мозга - блок программирования, регулирования и контроля над протеканием психической деятельности - включает моторные, премоторные и префронтальные отделы коры лобных отделов головного мозга. Кора лобных долей мозга занимает 24% поверхности больших полушарий. В лобной коре выделяют моторную кору (4, 6) и немоторную (9, 10, 11, 12, 46, 47). Эти области имеют различное строение и функции. Моторная лобная кора составляет ядерную зону двигательного анализатора.
Общая структурно-функциональная модель организации мозга, предложенная А.Р. Лурия, предполагает, что различные этапы произвольной, опосредованной, осознанной психической деятельности осуществляются с обязательным участием всех трех блоков мозга.
Согласно современным психологическим представлениям, каждая психическая деятельность имеет строго определенную структуру: она начинается с фазы мотивов, намерений, замыслов, которые затем превращаются в определенную программу деятельности, включающую «образ результата» и представления о способах реализации этой программы, а затем реализуется с помощью определенных операций. Завершается психическая деятельность этапом сличения полученных результатов ñ исходным «образом результата». В случае несоответствия полученных данных психическая деятельность продолжается до получения нужного результата. Поражение любого из трех блоков (или нарушение каких-либо отделов этих блоков) отражается на любой психической деятельности, так как приводит к нарушению соответствующей стадии или этапа ее реализации. Данная общая схема формирования мозга как субстрата сложных сознательных форм психической деятельности находит конкретное подтверждение при анализе различных нарушений высших психических функций, возникающих вследствие локальных поражений головного мозга.
Белое вещество полушарий
К белому веществу полушария относятся внутренняя капсула, свод и волокна, проходящие через спайки мозга, а также волокна, соединяющие в полушарии отдельные участки коры между собой, кору и базальные ядра.
Нервные волокна в полушарияхделят на три группы: ассоциативные, комиссуральные и проекционные.
Ассоциативные волокна связывают между собой участки одного и того же полушария. Короткие волокна соединяют соседние извилины, а длинные волокна – доли полушария.
Комиссуральные волокна связывают симметрично расположенные участки разных полушарий, образуя при этом спайки. К ним относятся: мозолистое тело, передняя спайка мозга и спайка свода.
Проекционные волокна связывают кору полушарий мозга с нижележащими отделами головного и спинного мозга. При этом выделяют восходящие и нисходящие волокна. Проекционные волокна, находящиеся ближе всего к коре, образуют лучистый венец. Все они затем сходятся во внутреннюю капсулу, представляющую собой слой белого вещества, расположенный между хвостатым ядром, чечевицеобразным ядром и зрительным бугром. На горизонтальном разрезе капсула имеет вид бумеранга, открытого в латеральную сторону.
Свод находится под мозолистым телом. В области средней трети мозолистого тела свод, раздваиваясь, дугообразно загибается впереди вниз. При этом образуются столбы свода, которые заканчиваются в мамиллярных телах гипоталамуса. Кроме того, раздваиваясь кзади, свод образует ножки свода, которые идут в нижний рог бокового желудочка. Между столбами свода и ножками свода натянута тонкая прозрачная перегородка.
Проводящие пути
Проводящие пути–это пучки нервных волокон, проходящих в определенных зонах белого вещества головного и спинного мозга, объединенных общностью морфологического строения и функции, и соединяющих между собой нервные центры. В ЦНС выделяют восходящие и нисходящие проводящие пути. Ввиду их большого количества мы рассмотрим только основные.
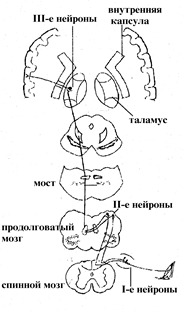
Путь сознательной проприоцептивной чувствительности
Восходящие пути.Путь сознательной проприоцептивной чувствительности (путь Голля и Бурдаха). Первые нейроны этого пути лежат в спинальном ганглии. Причем от 19 нижних ганглиев идут волокна, образующие пучок Голля, а от 12 верхних – пучок Бурдаха. Аксоны первых нейронов проходят в составе задних корешков и, не заходя в серое вещество задних рогов, проходят в задние канатики, поднимаются вверх и оканчиваются в продолговатом мозге в нежном ядре и клиновидном ядре, соответственно.
В этих ядрах лежат тела вторых нейронов. Их аксоны совершают перекрест и в составе медиальной петли проходят транзитом через мост, средний мозг и заканчиваются в латеральных ядрах таламуса. Здесь располагаются тела третьих нейронов. Их аксоны проходят через внутреннюю капсулу и оканчиваются в задней центральной извилине.
Дата добавления: 2015-08-08; просмотров: 1724;