Быстрый сон — это совершенно особое, по сравнению с медленным сном, состояние мозга.
Лишение животных парадоксального сна делает их возбудимыми, раздражительными. Это приводит впоследствии к компенсаторному увеличению продолжительности парадоксального сна и «вхождению» в него, минуя обычную промежуточную стадию медленноволнового сна.
Известная независимость быстрого и медленного сна подтверждается и различиями в их физиологическбй и химической природе. Установлено, что структуры, ответственные за медленный сон, находятся в каудальной части мозгового ствола, главным образом — в продолговатом мозге. Наличие сходных гипногенных структур было установлено также и в задней части моста.
В других исследованиях показано, что поведенческие и ЭЭГ-проявления парадоксальной стадии сна связаны с активацией структур в области моста. Это прежде всего зона голубого пятна,
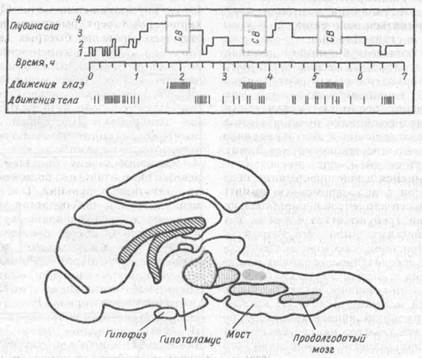
Рис.32. Механизмы организации сна (по Г.А. Адаму, 1983)
внизу — «центры» сна (гипногениые центры) в стволе мозга кошки: участки с вертикальной штриховкой — ядра, связанные со сном без сновидений; зачерненный участок соответствует расположению нейронов, контролирующих парадоксальный сон; вверху ритмическое чередование движений глаз и тела в течение семи часов сна, включающих три фазы быстрых движений глаз (СВ — сновидения)
а также ретикулярные и вестибулярные ядра. В стадии быстрого сна возникают в гиппокампе периоды тета-ритма, которые связывают с вегетативными сдвигами и движениями глаз (Т. Н. Ониани и др., 1970).
М. Жуве (1967) развивает гипотезу о моноаминергическлй регуляции стадий сна. По его представлениям медленновол новый сон регулируется серотонинергической системой ядер шва в продолговатом мозге (рис. 33, Б], ибо им установлена прямая зависимость сна от функциональной активности этих мозговых образований и общего уровня серотонина и его обмена. Катехоламинергические нейроны голубого пятна и ретикулярной формации среднего мозга выступают как антагонистическая система, ответственная за возникновение быстрого сна и состояния бодрствования (рис. 33, А, В).
Существуют указания на особую роль глиальных элементов во время сна. Н. Н. Демин и др. (1978) полагают, что одной из основных нейрохимических функций медленного сна являются пластические репарационные процессы, связанные с метаболизмом белков и РНК, причем главным образом в глиальных клетках определенных-структур мозга. Активные конформационные изменения в мембранах нейронов при парадоксальном сне предназначены для ликвидации так называемой усталости синаптического проведения и являются одной из причин своеобразия ЭЭГ при данном функциональном состоянии.
Ф Биологическое значение быстрого сна продолжает оставаться неразрешенной проблемой. Это видно даже из краткого перечня возможных фук-ций этого физиологического состояния (С. Хартман, 1967): 1) нейтрализа-
ция токсических веществ, накопившихся во время деятельности; 2) генерация квазисенсорных импульсов, стимулирующих развитие мозга; 3) восстановление функциональной организации рабочих механизмов мозга, угнетенных в течение медленного сна; 4) подготовка к переходу в состояние бодрствования; 5) освобождение от накопившейся необработанной информации; 6) передача сообщения из оперативной памяти в долгосрочную; 7) организация сновидений и текущей адаптации организма к условиям среды.
Сезонные ритмы поведения.Наряду
с циркадными ритмами поведение живых организмов подчинено также сезонным колебаниям окружающей среды, при которых изменяются условия питания, температуры, влажности, солнечного режима и т. д. Наиболее известные формы сезонного поведения проявляются в миграциях и кочевках, в явлениях зимней и летней спячки, а также в разнообразной деятельности по постройке гнезд и убежищ. Главными факторами при этом являются режим освещения и температура, а в качестве комплекса ответных' реакций — изменение гормональной активности, осуществляющей запуск репродуктивного цикла.
Сезонная периодика поведения оказывается чрезвычайно устойчивой. Так, например, было отмечено, что австралийский страус в заповеднике Аскания Нова откладывает яйца в условиях нашей зимы, несмотря на сильный мороз, прямо в снег в сезон, соответствующий лету в Австралии (М. М. За-вадовский, 1930).
Миграционная деятельность животных приурочена к определенным сезо1 нам года. Перелет птиц, миграция рыб, крупных млекопитающих связаны прежде всего с сезонными изменениями условий питания и характеризуются всеми чертами врожденного поведения, которому присущи некоторые общие черты, такие, как изменения стадных отношений, изменения в функции нейросекреторной системы, изменения характера терморегуляции, накопление жира в предмиграционный период.
Понижение температуры среды рас-
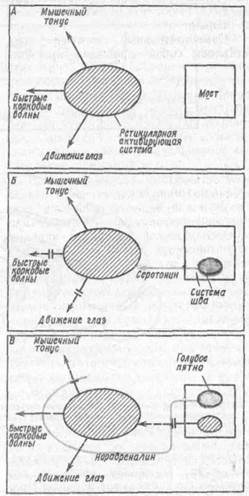
''пг: 31 Схематическое представление о пейрофи
;:•!'•..': "i?i\ri f! 1-ГЙрОХИМ!ГИ СНГ, (ПО ЭД ЖVIIС, 1067)
Дата добавления: 2015-08-04; просмотров: 1413;