Восприятие пространства и пространственная ориентация
Теории рефлекторного отражения пространства.Ориентация животного во времени и пространстве-осуществля-ется с помощью головного мозга с его внешними рецспторными аппаратами.
И. М. Сеченов особую роль в пространственном восприятии отводил двигательному аппарату и создаваемому им в головном мозгу «темному мышечному чувству». В непрерывной связи чувствования и движения И. М. Сеченов видел суть рефлекторного отражения пространства и времени. Особая роль мышечного аппарата обусловливается тем, что он является одновременно и рабочим органом и периферическим отделом «чувствующего снаряда». Другим орудием И. М. Сеченов считал те раздражения, которые поступают в головной мозг из внешней среды благодаря деятельности сенсорных органов, как сочетанную и координированную деятельность специальных форм чувствования между собой и с двигательными реакциями организма.
В. М. Бехтерев (1884, 1896) также рассматривал механизм отражения пространственных отношений как взаимосвязанную деятельность органов равновесия с внешней рецепцией и двигательным аппаратом. Существенное значение он придавал кожно-мышечному чувству.
А. А. Ухтомский (1954) отмечал, что конкретное восприятие совершается всегда в пространстве и времени нераздельно, в хронотопе. При зритель-
ной оценке предмета человек руководствуется не только зрительными и проприоцептивными {от мышц глаза) импульсами, но и одновременными рецепциями со слухового, вестибулярного и тактильно-проприоцсптивного аппаратов.
Экспериментально установлено (Э. Ш. Айрапетьянц и А. С. Батуев, 1963), что пространственный анализ обеспечивается комплексом динамически увязанных между собой анализаторов и среди них в особенности — зрительного, вестибулярного, кожного и мышечного. Интеграция комплекса и его реинтеграция в случае повреждения обеспечиваются при непременном участии высших ассоциативных систем мозга.
Исключительность пространственно-различительной функции какой-либо одной сенсорной системы противоречит экспериментальным данным (Б. Г. Ананьев, 1961).
Концепции И. С. Бериташвили о пространственной ориентации и А. А. Ухтомского об интегральном образе.И. С. Бериташвили предложил концепцию психонервного поведения, которое регулируется целостным «представлением» о внешней среде или образом той внешней среды, в которой находится и действует животное. В понятие образа среды входят все объекты внешнего мира, которые имеют связь с биологически мотивированным состоянием животного. На основе первого знакомства со средой животное вырабатывает определенные «представления» («гипотезы» по Я. Кречевскому) об ее организации и использует их в форме поведенческих тактик взаимодействия со средой с целью проверки, корректировки и дальнейшего их усовершенствования. Это составляет тот уровень эвристической деятельности мозга в новых ситуациях, который прогрессивно развивается в ходе эволюции и на его основе формируются жесткие условно-рефлекторные программы автоматизированного поведения (Т. А. Натишвили, 1987).
А. А. Ухтомский (1924) разработал концепцию об интегральном образе.
Простое ощущение, отмечал А. А. Ухтомский, есть в сущности абстракция, более или менее полезная аналитическая фикция, тогда как реальный и живой опыт имеет дело с интегральными образами. Всякий интегральный образ является продуктом пережитой доминанты.
Итак, имея биологическую потребность в пище, животные и человек, однократно столкнувшись с возможностью ее получения, сразу же формируют комплексный образ местонахождения пищи (И. С. Бериташвили). По А. А. Ухтомскому, это первая стадия развития доминанты, когда она «привлекает» к себе самые разнообразные внешние рецепции. По И. П. Павлову,— это стадия генерализации условного рефлекса, которая протекает по механизмам доминанты. Генерализация условного рефлекса создает обобщенный пространственный образ среды, который при наличии высокого уровня мотивации фиксируется в памяти, т. е. формирует конкретное представление.
Образное поведение — это самый первый этап индивидуального обучения. Дальнейшее развитие поведения может происходить двумя путями. Либо организм из множества пространственных факторов среды извлекает такие, которые с наибольшей вероятностью приведут к удовлетворению текущей биологической потребности. В этом случае формируется прочная временная связь между ограниченным набором сигналов и соответствующей поведенческой реакцией — вырабатываются стабильные специализированные условные рефлексы и организм переходит на режим автоматического управления.
Но допустим и другой путь развития индивидуального поведения, детерми-нантой в котором остается тот же образ, но прочно зафисированный в энграмме памяти. Что побуждает организм формировать поведение по этому второму пути, который представляет собой стадию закрепившейся доминанты? Во-первых, высокий уровень мотивации, а во-вторых, низкая вероятность ее удовлетворения. При однократном подкреплении пищей очень голодного

Рис 27 Пары изображений пространственных обьектов, предъявляемые испытуемым в опытах по мысленному вращению объектов (по Т. А. Натишвилли, 1987)
верхняя пара — идентичные объекты, повернутые относительно друг друга в плоскости экрана; средняя — идентичные объекты, повернутые относительно друг друга в трех измерениях; нижняя зеркальные конфигурации одного и того же объекта. Испытуемые должны установить, какие пары являются идентичными изображениями одного и того же объекта, а в каких использованы зеркальные конфигурации данного объекта
животного организм не в состоянии оцепить вероятность повторного получения пищи в аналогичной ситуации. Однако в связи с тем, что естественная среда чаще всего создает именно такую ситуацию, т. е. не может гарантировать высокую вероятность удовлетворения потребности в постоянно меняющейся обстановке, естественный отбор жестко зафиксировал в эволюции свойство не только воспринимать комплексное воздействие окружающей среды, но и быстро фиксировать его в виде целостного образа.
В дальнейшем будет достаточно лишь одного компонента, чтобы репродуцировать целый образ и направить поведение сообразно этому образу.
Эти два крайних механизма поведения (которые моделируются в лабораторных условиях) подразумевают в естественной среде и наличие соответствующих механизмов. Они строятся на сложных взаимоотношениях обоих детерминант: как вероятностной программы (т. е. интегрального образа), создающей вектор поведению, так и автоматизированных условных рефлексах, обеспечивающих адаптивный характер реализации этой программы в знакомой пространственной ситуации.
Феномен мысленного вращения объектов.В психологических исследованиях на человеке было установлено (Шепард, 1924) наличие образного представления внешнего пространства. В основу был положен феномен мысленного вращения. Испытуемым предъявлялись пары рисунков трехмерных объектов, различающиеся своей ориентацией (рис. 27). Испытуемые должны были решить, являются рисунки идентичными или зеркальными копиями. Оказалось, что время реакции возрастает с увеличением угла, на который один из элементов пары повернут относительно другого. Испытуемый производит мысленное вращение одного из элементов пары до геометрического его совмещения со вторым и на основании этого заключает об их идентичности или нсидснтичности. Причем в процессе мысленного вращения сохраняется значительная доля образной информации об объекте. Такое образное представление реальности можно также назвать аналоговым представлением (Т. А. Натишвили, 1987). ,
Есть основаниия предполагать, что аналогичные мозговые процессы могут осуществляться и высшими млекопитающими. Шепард приводит следующее наблюдение. Овчарка доставала длинную палку, переброшенную через забор, в котором отсутствовала вертикальная доска. Собака проникла в щель, схватила в зубы палку и, повернувшись, двинулась назад к отверстию. В

Рис. 28, Схема двенадцатилучевого радиально-симметричного лабиринта (по К. А. Ря бинской, 1987):
А — вертикальный разрез; Б -- вид сверху, цифры — номера коридоров
последний момент она вдруг остановилась и после некоторой паузы повернула голову па 90°. Держа палку в таком положении, она свободно пролезла через щель в заборе.
Концепция когнитивных карт.Под когнитивными (познавательными) картами понимается процесс, благодаря которому организм приобретает некое подобие топологической карты той местности, в которой он обитает. Когнитивная карта — это динамический образ, способный к изменениям и уточнениям на основании информации об изменении среды либо при изменении местонахождения в ней самого субъекта (Е. Толмен, 1948).
Д. С. Олтон (1979) разработал тесты, с помощью которых было показано, что крысы способны запоминать точное местонахождение пищи в пространстве и направление к нему независимо от исходной стартовой позиции.
Один из широко распространенных приемов для изучении когнитивного картирования состоит в использовании
12-лучевого радиального лабиринта (рис. 28). Коридоры оборудованы входными и выходными дверцами, открывающимися в центробежном направлении. В конце каждого из них помешается пища. Каждый выбор начинается с центра платформы, затем животное идет в один из коридоров, опускается вниз, и цикл вновь повторяется. Повторное посещение коридора не сопровождается пищевым вознаграждением. Получение наибольшего количества пищи достигается одноразовым посещением всех 12 коридоров, т. е. поведением, строго соответствующим когнитивной пространственной карте. Наблюдение за поведением животных в таком лабиринте позволило установить определенные закономерности, которые получили название стратегий поведения, или гипотез. Концепция поведенческих стратегий или гипотез противостоит идее о случайном переборе животным различных реакций (метод проб и ошибок). Я. Кречевский (1932) допускал, что жи-
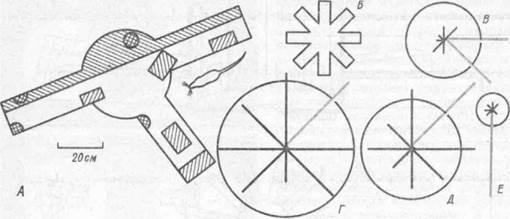
Рис.29. Поля нейронов в гиппокампе у крыс (по Л.И. Францевичу, 1986):
А — участки лабиринта, в котором активируются определенные нейроны (заштрихованы) (О. Кифи, 1976); Б — план восьмилучевого лабиринта; В — Е — диаграммы активности отдельных нейронов в лучах лабиринта: окружность — уровень фпновой активности (Дж. Олтон, 1978)
вотное не поступает наугад, а формирует и проверяет серии «гипотез» для решения задачи.
Наиболее обоснованно представление о существовании трех основных стратегий решения животными задач пространственной ориентации. Первая — использование когнитивных карт местности, вторая — поиск опорных внешних ориентиров и третья - поиск на основе последовательности собственных двигательных реакций. Первые две стратегии являются аллоцент-рическими (стратегии места), основанными на пространственной топологии среды. Третья стратегия получила название эгоцентрической (стратегия ответа), основанная на отношении между элементами пространства и положением и перемещением тела животного в этом пространстве. Если подобные стратегии и существуют, то они постоянно сменяют друг друга и часто выступают в смешанной форме даже в процессе реализации одной поведенческой задачи.
Анализ поведения крыс в 12-лучевом лабиринте выявил закономерности, именуемые поведенческими тактиками (Е. А. Рябинская, 1987). Так, например, крысы посещают коридор, более удаленный от коридора, который они посещали до этого. Кроме того, крысы чаще выбирают коридор, расположенный
справа или слева по отношению к коридору, который они только что покинули.
Э. Хониг (1978) предлагает для анализа поведения в сложных пространственных условиях выделять рабочую память (запоминание информации, необходимой только в пределах одного опыта) и референтную память (запоминание информации, регулярно повторяющейся во всех опытах). И тот и другой виды пространственной памяти подразумевают участие обоих видов стратегий поведения, аллоцентрической и эгоцентрической. Участие собственно двигательного компонента весьма велико, хотя животные способны прогнозировать возможные последствия самых различных движений, даже таких, которые, имеются лишь «в плане», но еще никогда не выполнялись (Т. А. На-тишвили, 1987).
Нейрофизиологические корреляты пространственного восприятия.Специальными опытами на крысах было показано, что их поведение в радиальном лабиринте зависит от целостности гиппокампальной системы. Ее нарушение не приводит к дефициту аллоцентрической пространственной локализации, но приводит к дефициту оперативной памяти, по которой отбираются коридоры лабиринта.
Заднетеменная кора представляет
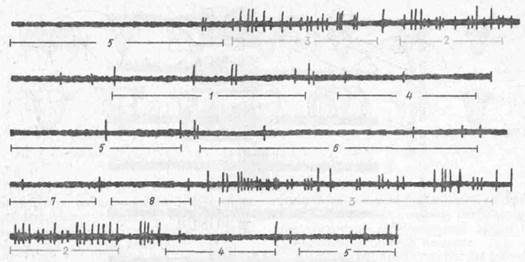
Рис. 30. Активность нейрона дорзального гиппокампа крысы при пробежках гю коридорам ради лабиринта (но Дж. Олтону, 1977}
(Осциллографическая записьактивности одиночной клетки, регистрируемой металлическим микроэлектродом. Уровень активности клетки строго коррелирует с местонахождением животного в пространстве. Импульсация клетки низка при нахождении животного в коридорах 1, 4. 5, 6, 7 и 8, высока при местонахождении животного в коридорах 2 и 3. Тем самым рецептивное поле данной клетки разделяет весь радиальный лабиринт на небольшое on-поле и значительно большее оff-поле.)
собой часть нервного аппарата, специализированного для анализа и запоминания взаимного пространственного расположения предметов.
Интересная картина выявляется при оценке активности нейронов гиппокампа при ориентации крыс в коридорах радиального лабиринта (О'Кифи, 1976; Олтон, 1987). Отдельные нейроны гиппокампа возбуждались при нахождении животного в определенных лучах лабиринта (рис. 29). Некоторые нейроны возбуждались особенно сильно, когда крыса находилась в специфическом участке и в определенной ориентации относительно лабиринта (рис. 30). Поворот лабиринта в системе внешних ориентиров не менял специфичности «нейронов места» в отношении идентифицируемых участков лабиринта. На активность нейрона не влияло ни отвлечение внимания животного, ни голодное или сытое его состояние, ни тип поведения животного в этом участке лабиринта. Такие свойства клеток формируются очень быстро — не позднее, чем через 10 мин после попадания животного в новую обстановку. И раз сформировавшись, такие клетки оказывают-
ся весьма стабильными в отношении собственного пространственного рецептивного поля. По-видимому, нейроны гиппокампа участвуют в узнавании места и соотнесении его с координатами какой-то пространственной карты на основе аллоцентрической системы пространственной локализации, т. е. опять же в реализации оперативной памяти.
В исследовании на овцах с регистрацией активности нейронов нижневисочной коры было обнаружено восприятие зрительного изображения головы представителей своего же вида, но снабженных рогами (К. Кендрик, Б. Балд-вин, 1987) (рис. 31). Из этих опытов делается вывод об участии нижневисочной коры в восприятии целостных пространственных образов, имеющих отношение к социальному поведению животных.
Предполагают существование у млекопитающих мозговой системы, выполняющей функцию пространственной ориентации (Л. И. Францевич, 1986); нижневисочная кора — узнавание зрительных ориентиров, гиппокамп — узнавание топологии ориентиров в пространстве, теменная кора — установление
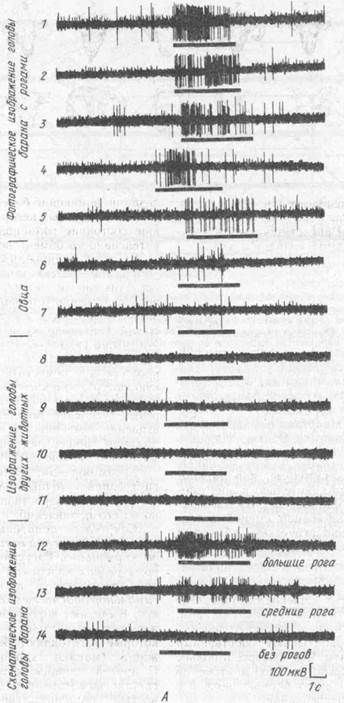
Рис. 31. Ответы нижневисочной области коры у овцы (А), отвечающие на предъявления изображении
лица (Б)(по.К.М. Кендрику и Б.А.Болдвину, 1987).
(Приведены ответы одиночной клетки на последовательное предъявление разных стимулов Момент предъявления обозначен черной горизонтальной стрелкой. Видно, что нейрон реагирует речким усилением импульсной активности на изображения, в которых присутствуют рога. Величина ответа зависит от величины рогов)

Рис. 31 Продолжение.
взаимного расположения ориентиров, префронтальная кора или ее аналоги — запоминание и расчет действий в установленной системе координат тела.
Дата добавления: 2015-08-04; просмотров: 3360;