НОРМА РЕАКЦИИ И ДИАПАЗОН РЕАКЦИИ
Еще два понятия, овладение которыми чрезвычайно важно для правильного понимания отношений между генотипом и фенотипом, — это «норма реакции» и «диапазон реакции». Семантические поля этих
двух понятий близки, однако между ними существуют весьма значимые различия. Описывая далее взаимоотношения между этими понятиями, мы сначала остановимся на том, что является для них общим, а затем — на их отличительных признаках.
Общее в понятиях нормы и диапазона реакции заключается в следующем. Нормой (диапазоном) реакции данного генотипа называется система, описывающая множество фенотипов, существование которых потенциально возможно в том случае, если данный генотип будет находиться во взаимодействии с определенными средами. Понятия и нормы, и диапазона реакции предполагают, что каждый генотип ассоциируется с определенным, характерным для него, рядом фенотипов, формирующихся в разных средах. Упрощенно понятие нормы (диапазона) реакции можно представить следующим образом:
Различия в понятиях нормы и диапазона реакции состоят в следующем. Рассмотрим гипотетический пример, касающийся фенотипи-ческого признака, который отражает какие-то специфические способности. Предположим, существует 4 генотипа (1, 2, 3, 4), и все эти генотипы могут быть одновременно помещены в разные типы сред, отличающиеся друг от друга по уровню разнообразия и обогащеннос-ти. Схематически эта ситуация отражена на рис. \.5а. По оси абсцисс отложены «уровни» среды, а по оси ординат — условные фенотипи-ческие значения. Согласно рисунку, в обедненной среде разброс фе-нотипических значений относительно мал, и четыре генотипа прояв-
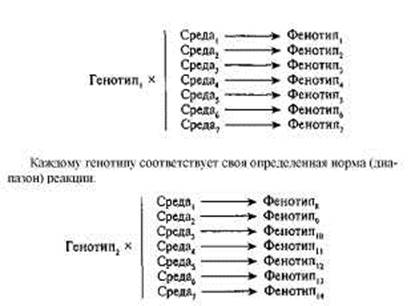
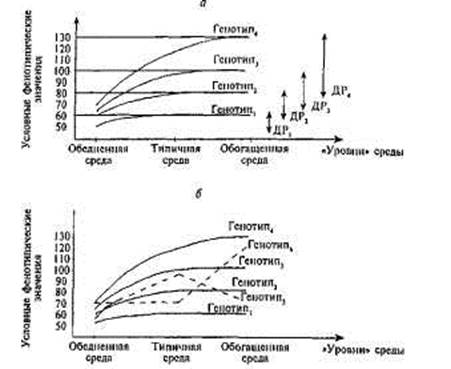
Рис. 1.5. Схематическое изображение зависимости фенотипических значений от типов генотипа и среды, в которой данный генотип развивается.
а— Схематическая иллюстрация понятия «диапазон реакции» (ДР). Диапазон реакции обозначает разницу между фенотипическими значениями определенного генотипа в разных типах сред [по: Gottesman, 1963]. б — Схематическая иллюстрация понятия «норма реакции». Один и тот же генотип (Генотип5 и Генотип6) в разной среде дает разные фенотипы.
ляются в фенотипах, мало отличающихся друг от друга. Разброс фенотипических значений существенно возрастает в типичной среде и достигает максимума в среде обогащенной. Разница между значениями данного генотипа в обедненной и обогащенной средах называется диапазоном реакции этого генотипа.
Обратите внимание: понятие диапазона реакции подразумевает сохранение рангов фенотипических значений генотипов в разных сре-довых условиях. Например, Генотип1 ассоциируется с низкими фенотипическими значениями и в обедненной, и в обогащенной средах, в то время как Генотип4 является наиболее «процветающим» в любой среде. Соответственно, диапазон реакции Генотипа1 — наименьший,
а диапазон реакции Генотипа4 — наибольший. Иными словами, основным допущением при интерпретации понятия «диапазон реакции» служит следующее предположение: существующие генотипы отличаются друг от друга таким образом, что фенотипические преимущества каждого из этих генотипов постоянны, а фенотипические различия, ассоциируемые с каждым из генотипов, становятся все более заметны по мере того, как среда становится все более благоприятной для развития данного фенотипического признака. Если взять в качестве примера математические способности, то носители Генотипа4 будут демонстрировать наивысшие значения как в обедненной, так и в обогащенной среде, причем чем благоприятнее среда, тем выше уровень математических достижений. Напротив, носители Генотипа1 будут иметь наименьшие фенотипические значения в любой среде, а фенотипические изменения, характеризующие этот фенотип при переходе из одних средовых условий в другие, будут незначительны.
Добавим к изучаемым нами генотипам два новых — Генотип5 и Генотип6 (рис. 1.5б). Оказывается, что поведение этих двух генотипов в разных средах не соответствует ожиданиям о сохранении ранговых мест фенотипических выражений разных генотипов в варьирующих средовых условиях. Как показано на рис. 1.5б, максимальное феноти-пическое значение Генотипа5 наблюдается в типичной среде, в то время как обогащенная среда не является благоприятной для этого генотипа — его фенотипическое значение уменьшается. В качестве возможной иллюстрации данного феномена может быть использован хорошо известный из психологии развития факт: излишняя когнитивная стимуляция многих (но не всех) младенцев часто приводит не к оптимизации, а к расстройству их познавательной деятельности.
Генотип6, напротив, на переход от обедненной к типичной среде никак не реагирует, его фенотипическое значение остается неизменным. Однако ситуация существенно меняется при изменении средовых условий на обогащенные: фенотипическое значение Генотипа6 резко и линейно возрастает. Примером подобной ситуации может служить развитие музыкальных способностей, поскольку ребенок, основываясь на своих природных задатках, должен овладеть мастерством, для обучения которому ему необходимо находиться в обогащенной среде, в то время как и обедненная, и типичная среды таких условий не дают.
Таким образом, несколько упрощая ситуацию, можно сказать, что понятие нормы реакции — более общее понятие, поскольку, используя его, исследователь не должен делать никаких предположений о сохранении рангов фенотипов в разных средах. Для понятия же диапазона реакции допущение об определенном ранговом порядке фенотипов (и, соответственно, генотипов) в контексте разных средовых условий является критическим. В силу большей широты понятия нормы реакции далее в учебнике будет использоваться именно это понятие.
5-1432 65
Сегодня мы не располагаем аналитическими средствами, которые позволили бы нам предположить, что произойдет с индивидуумом, являющимся носителем определенного генотипа, если он будет помещен в среду, отличающуюся от любой предыдущей; поэтому определение нормы реакции — задача экспериментальная. Каким образом норма реакции генотипа определяется в эксперименте? С этой целью генетически одинаковые организмы помещаются в разные среды, а фенотипы, развивающиеся в результате взаимодействия генотипа и различных сред, тщательно измеряются и описываются. В процессе генетического экспериментирования исследователь старается выделить для анализа ограниченное количество генов, детерминирующих определенные признаки. Исследователь также старается застраховаться от случайных влияний среды, работая в максимально контролируемых условиях. Понятно, что этот тип эксперимента возможен только с растениями или животными.
Задача определения набора сред, приводящих к проявлению одного и того же генотипа в разных фенотипах у человека (или определение нормы реакции данного генотипа), чрезвычайно сложна. Для изучения нормы реакции необходимо некоторое количество генетически идентичных особей, т.е. необходимы группы идентичных близнецов (пятерняшек, шестерняшек или, того лучше, двадцатерняшек), родители которых согласились бы разлучить детей при рождении и растить их в разных средовых условиях. Биологически задача создания генетически идентичных человеческих организмов очень трудна, но осуществима; с точки же зрения этики такой эксперимент в цивилизованном обществе принципиально невозможен.
Не менее сложна задача определения параметров среды, существенных для развития изучаемого признака человека. Например, что является оптимальной средой для формирования интеллектуальной активности ребенка? Или уже: что в семейной среде стимулирует интеллектуальную активность ребенка — количество книг, наличие компьютера или чтение сказок перед сном? Решая данные задачи, психогенетика находится в прямом и непосредственном контакте с психологией и, более того, зависит от нее, поскольку именно из психологии психогенетика должна получать сведения о том, какие конкретные средовые условия существенны для развития того или иного психологического признака.
«Норма реакции» является понятием по своей природе интерак-ционистским, т.е. подчеркивающим идею взаимодействия вовлеченных в развитие факторов генотипа и среды. Конкретный фенотип представляет собой реализацию конкретного генотипа в конкретных средовых условиях в соответствии с его нормой реакции, и процесс этого взаимодействия чрезвычайно сложен. Любое искусственное расчленение и квалификация генотипических и средовых влияний на формирующийся организм является его упрощением, и это необходимо помнить при интерпретации психогенетических данных.
5. ДВА ПОДХОДА К АНАЛИЗУ СВЯЗЕЙ МЕЖДУ ГЕНОТИПОМ И ФЕНОТИПОМ
Между генотипом и фенотипом нет неопосредованной зависимости. Между геном и первичным биохимическим проявлением его действия (например, синтезом какого-либо белка), с одной стороны, и влиянием этого гена на поведение — с другой, прямого соответствия не существует. Влияние генов на поведение имеет опосредованный характер. В той мере, в какой индивидуальные различия в психике и поведении передаются по наследству, они представляют собой результат сложнейших биохимических процессов. Непосредственное биохимическое проявление гена и его влияние на психологические особенности разделены «горным хребтом» промежуточных биомолекулярных событий.
Для изучения зависимости между геном (или генотипом) и поведением (или фенотипом) исследователи располагают двумя подходами. Разница этих подходов определяется начальной точкой движения: первый подход предполагает движение от фенотипа к генотипу, второй — от генотипа к фенотипу. Отправляясь от наблюдаемого (от фенотипа), исследователь должен прежде всего удостовериться в том, что анализируемый признак действительно подвержен влиянию со стороны данного гена, и только затем переходить к изучению последнего. В рамках этого подхода сначала изучаются законы передачи анализируемого признака по наследству, затем картируют* ген, детерминирующий развитие этого признака, а потом изучают генный продукт (белок).
Второй подход предполагает противоположное направление движения — от гена (генов) к фенотипу. Данная аналитическая стратегия заключается в локализации изучаемого гена, определении его структуры и описании его нуклеотидов. Знание последовательности нукле-отидов на участке ДНК, функция которого неизвестна, позволяет сделать вывод о последовательности аминокислот в белке, за синтез которого этот участок отвечает. Зная такую последовательность, можно синтезировать искомый белок, а затем ввести его животному с целью изучения его функций. Инъекция «чужого» белка вызывает образование у животного специфических антител, которые дают возможность выяснить, в клетках какого типа производится изучаемый белок и какова его функция. Более того, зная нуклеотидную структуру гена, ответственного за производство изучаемого белка, исследователь может вызвать искусственные мутации гена. Изменив структуру данного белка, можно изучать изменения в фенотипе, вызываемые такими мутациями.
Психогенетика как наука, развивающаяся на стыке генетики и психологии, характеризуется двойственностью своих исследовательс-
* Картирование — составление генетических карт хромосом.
5* 67
ких задач: они пересекаются с задачами собственно генетическими и собственно психологическими. Конечной целью генетического исследования человеческого организма, общей с целями генетических исследований других живых организмов, является идентификация гена (генов), ответственного за формирование тех или иных поведенческих признаков, его положения на хромосомной карте и описание гена и его продуктов. Отсутствие продуктов этого гена — носителя определенного поведенческого признака — в организме человека или обнаружение корреляции между мутацией гена и анализируемым признаком служат свидетельством того, что найденный ген вовлечен в формирование и(или) проявление анализируемого признака.
После того как ген картирован и его продукт описан, изучение белка, синтез которого контролируется исследуемым геном, может привести к пониманию физиологического механизма изучаемого признака. Исследование физиологического механизма признака, в свою очередь, может помочь разработке профилактических программ (как биологических, так и небиологических), целью которых является уменьшение или полная остановка неблагоприятного влияния белков, синтезируемых в результате вредоносных мутаций исследуемого гена. В конце концов, поняв систему, в которую вовлечен изучаемый ген, исследователи, возможно, смогут разработать программы, позволяющие заменять вредоносные аллели-мутанты новыми, «здоровыми» вариантами гена.
Конечной целью психологического исследования является понимание этиологии и структуры анализируемого признака, обнаружение факторов, влияющих на его индивидуальное развитие, и тех характеристик среды (культурной, социальной, групповой, семейной или индивидуальной), которые позволяют влиять на развитие, а также макро- и микрофункционирование данного признака.
Психогенетические исследования подчиняются обеим целям, изучая генотип и среду в их непрерывном взаимодействии, объединяя в себе методологию и инструментарий обеих наук.
* * *
Изменчивость, межиндивидуальная вариативность — неизбежная форма существования живых организмов. Она формируется в результате взаимодействия наследственных и средовых факторов, комбинация которых уникальна для каждого живого организма.
Общие закономерности наследования признаков систематизированы в рамках хромосомной теории наследственности, центральные понятия которой — «хромосома», «генотип», «ген» и «аллель».
Продуктом реализации данного генотипа в данной среде является фенотип — наблюдаемые морфологические, физиологические, пси-
хологические характеристики организма. Фенотипы не наследуются, а формируются в течение жизни в результате взаимодействия генотипа и среды. Одним из центральных понятий при описании этого взаимодействия служит понятие «норма (диапазон) реакции».
Глава II
КЛАССИЧЕСКИЕ ЗАКОНЫ Г.МЕНДЕЛЯ
1. ГЕНИАЛЬНОЕ ПРЕДВИДЕНИЕ ИЛИ ТВОРЧЕСКАЯ УДАЧА?
Основные законы наследуемости были описаны более века назад чешским монахом Грегором Менделем (1822-1884), преподававшим физику и естественную историю в средней школе г. Брюнна (г. Брно). Мендель занимался селекционированием гороха, и именно гороху, научной удаче и строгости опытов Менделя мы обязаны открытием основных законов наследуемости*: закона единообразия гибридов первого поколения, закона расщепления и закона независимого комбинирования.
Г. Мендель не был пионером в области изучения результатов скрещивания растений. Такие эксперименты проводились и до него, с той лишь разницей, что скрещивались растения разных видов. Потомки подобного скрещивания (поколение F1) были стерильны, и, следовательно, оплодотворения и развития гибридов второго поколения (при описании селекционных экспериментов второе поколение обозначается F2) не происходило. Другой особенностью доменделевских работ было то, что большинство признаков, исследуемых в разных экспериментах по скрещиванию, были сложны как по типу наследования, так и с точки зрения их фенотипического выражения.
Гениальность (или удача?) Менделя заключалась в том, что в своих экспериментах он не повторил ошибок предшественников. Как писала английская исследовательница Ш. Ауэрбах, «успех работы Менделя по сравнению с исследованиями его предшественников объясняется тем, что он обладал двумя существенными качествами, необходимыми для ученого: способностью задавать природе нужный вопрос и способностью правильно истолковывать ответ природы» [9]. Во-первых, в качестве экспериментальных растений Мен-
* Надо сказать, что некоторые исследователи выделяют не три, а два закона Менделя. Например, в руководстве «Генетика человека» Ф. Фогеля и А. Мотульс-ки (рус. изд. — 1989 г.) излагаются три закона, а в книге Л. Эрман и П. Парсонса «Генетика поведения и эволюция» (рус. изд. — 1984 г.) — два. При этом некоторые ученые объединяют первый и второй законы, считая, что первый закон является частью второго и описывает генотипы и фенотипы потомков первого поколения (F1). Другие исследователи объединяют в один второй и третий законы, полагая, что «закон независимого комбинирования» есть в сущности «закон независимости расщепления», протекающего одновременно по разным парам аллелей. Однако в отечественной литературе речь идет обычно о трех законах Менделя. Эту точку зрения принимаем и мы.
хологические характеристики организма. Фенотипы не наследуются, а формируются в течение жизни в результате взаимодействия генотипа и среды. Одним из центральных понятий при описании этого взаимодействия служит понятие «норма (диапазон) реакции».
Глава II
КЛАССИЧЕСКИЕ ЗАКОНЫ Г.МЕНДЕЛЯ
1. ГЕНИАЛЬНОЕ ПРЕДВИДЕНИЕ ИЛИ ТВОРЧЕСКАЯ УДАЧА?
Основные законы наследуемости были описаны более века назад чешским монахом Грегором Менделем (1822-1884), преподававшим физику и естественную историю в средней школе г. Брюнна (г. Брно). Мендель занимался селекционированием гороха, и именно гороху, научной удаче и строгости опытов Менделя мы обязаны открытием основных законов наследуемости*: закона единообразия гибридов первого поколения, закона расщепления и закона независимого комбинирования.
Г. Мендель не был пионером в области изучения результатов скрещивания растений. Такие эксперименты проводились и до него, с той лишь разницей, что скрещивались растения разных видов. Потомки подобного скрещивания (поколение F1) были стерильны, и, следовательно, оплодотворения и развития гибридов второго поколения (при описании селекционных экспериментов второе поколение обозначается F2) не происходило. Другой особенностью доменделевских работ было то, что большинство признаков, исследуемых в разных экспериментах по скрещиванию, были сложны как по типу наследования, так и с точки зрения их фенотипического выражения.
Гениальность (или удача?) Менделя заключалась в том, что в своих экспериментах он не повторил ошибок предшественников. Как писала английская исследовательница Ш. Ауэрбах, «успех работы Менделя по сравнению с исследованиями его предшественников объясняется тем, что он обладал двумя существенными качествами, необходимыми для ученого: способностью задавать природе нужный вопрос и способностью правильно истолковывать ответ природы» [9]. Во-первых, в качестве экспериментальных растений Мен-
* Надо сказать, что некоторые исследователи выделяют не три, а два закона Менделя. Например, в руководстве «Генетика человека» Ф. Фогеля и А. Мотульс-ки (рус. изд. — 1989 г.) излагаются три закона, а в книге Л. Эрман и П. Парсонса «Генетика поведения и эволюция» (рус. изд. — 1984 г.) — два. При этом некоторые ученые объединяют первый и второй законы, считая, что первый закон является частью второго и описывает генотипы и фенотипы потомков первого поколения (F1). Другие исследователи объединяют в один второй и третий законы, полагая, что «закон независимого комбинирования» есть в сущности «закон независимости расщепления», протекающего одновременно по разным парам аллелей. Однако в отечественной литературе речь идет обычно о трех законах Менделя. Эту точку зрения принимаем и мы.
дель использовал разные сорта декоративного гороха внутри одного рода Pisum. Поэтому растения, развившиеся в результате подобного скрещивания, были способны к воспроизводству. Во-вторых, в качестве экспериментальных признаков Мендель выбрал простые качественные признаки типа «или/ или» (например, кожура горошины может быть либо гладкой, либо сморщенной), которые, как потом выяснилось, контролируются одним геном. В-третьих, подлинная удача (или гениальное предвидение?) Менделя заключалось в том, что выбранные им признаки контролировались генами, содержавшими истинно доминантные аллели, И наконец, интуиция подсказала Менделю, что все категории семян всех гибридных поколений следует точно, вплоть до последней горошины, пересчитывать, не ограничиваясь общими утверждениями, суммирующими только наиболее характерные результаты (скажем, таких-то семян больше, чем таких-то).
Мендель экспериментировал с 22 разновидностями гороха, отличавшимися друг от друга по 7 признакам (цвет, текстура семян). Свою работу Мендель вел восемь лет, изучил 10 000 растений гороха. Все формы гороха, которые он исследовал, были представителями чистых линий; результаты скрещивания таких растений между собой всегда были одинаковы. Результаты работы Мендель привел в статье 1865 г., которая стала краеугольным камнем генетики. Трудно сказать, что заслуживает большего восхищения в нем и его работе — строгость проведения экспериментов, четкость изложения результатов, совершенное знание экспериментального материала или знание работ его предшественников.
Коллеги и современники Менделя не смогли оценить важности сделанных им выводов. По свидетельству А.Е. Гайсиновича [34], до конца XIX в, ее цитировали всего пять раз, и только один ученый — русский ботаник И.О. Шмальгаузен — оценил всю важность этой работы. Однако в начале XX столетия законы, открытые им, были переоткрыты практически одновременно и независимо друг от друга учеными К. Корренсом, Э. Чермаком и К. де Фризом. Значимость этих открытий сразу стала очевидна научному сообществу начала 1900-х годов; их признание было связано с определенными успехами цитологии и формированием гипотезы ядерной наследственности*.
2. ЗАКОН ЕДИНООБРАЗИЯ ГИБРИДОВ
ПЕРВОГО ПОКОЛЕНИЯ (ПЕРВЫЙ ЗАКОН МЕНДЕЛЯ)
Данный закон утверждает, что скрещивание особей, различающихся по данному признаку (гомозиготных по разным аллелям), дает генетически однородное потомство (поколение F1), все особи которого гетерозиготны. Все гибриды F1 могут иметь при этом либо фенотип одного из родителей (полное доминирование), как в опытах Менделя, либо, как было обнаружено позднее, промежуточный фенотип (неполное доминирование). В дальнейшем выяснилось, что гибриды первого поколения F1, могут проявить признаки обоих родителей (кодо-минирование). Этот закон основан на том, что при скрещивании двух гомозиготных по разным аллелям форм (АА и aа) все их потомки одинаковы по генотипу (гетерозиготны — Аа), а значит, и по фенотипу.
* Интересующимся историей генетики можно посоветовать прекрасное изложение ее в книге А.Е. Гайсиновича «Зарождение и развитие генетики» (М., 1988).
3. ЗАКОН РАСЩЕПЛЕНИЯ (ВТОРОЙ ЗАКОН МЕНДЕЛЯ)
Этот закон называют законом (независимого) расщепления. Суть его состоит в следующем. Когда у организма, гетерозиготного по исследуемому признаку, формируются половые клетки — гаметы, то одна их половина несет один аллель данного гена, а вторая — другой. Поэтому при скрещивании таких гибридов F1 между собой среди гибридов второго поколения F2 в определенных соотношениях появляются особи с фенотипами как исходных родительских форм, так и F1
В основе этого закона лежит закономерное поведение пары гомологичных хромосом (с аллелями А и а), которое обеспечивает образование у гибридов F1 гамет двух типов, в результате чего среди гибридов F2 выявляются особи трех возможных генотипов в соотношении 1АА : 2Аа : 1аа. Иными словами, «внуки» исходных форм - двух гомозигот, фенотипически отличных друг от друга, дают расщепление по фенотипу в соответствии со вторым законом Менделя.
Однако это соотношение может меняться в зависимости от типа наследования. Так, в случае полного доминирования выделяются 75% особей с доминантным и 25% с рецессивным признаком, т.е. два фенотипа в отношении 3:1. При неполном доминировании и кодомини-ровании 50% гибридов второго поколения (F2) имеют фенотип гибридов первого поколения и по 25% — фенотипы исходных родительских форм, т.е. наблюдается расщепление 1:2:1.
Приведем некоторые примеры этих типов наследования.
ДОМИНАНТНОЕ НАСЛЕДОВАНИЕ: БОЛЕЗНЬ ГЕНТИНГТОНА (ХОРЕЯ ГЕНТННГТОНА)
Хорея Гентингтона (ХГ) — дегенеративное заболевание нервных клеток в базальных структурах переднего мозга. Оно начинается с изменений личности больного и сопровождается прогрессирующей забывчивостью, слабоумием и появлением непроизвольных движений. Обычно заболевание диагностируется в зрелом возрасте (45-60 лет), и в течение последующих 15-20 лет пациент полностью теряет контроль над моторикой и когнитивной сферой. Способ лечения этого заболевания пока неизвестен. Частота встречаемости ХГ составляет примерно 1 на 20 000 человек, т.е. примерно четверть миллиона человек на земном шаре сегодня больны или в скором времени заболеют ХГ.
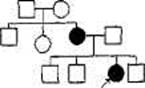
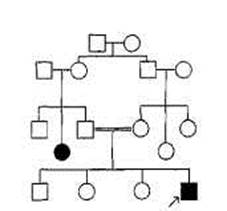
При изучении родственников больных ХГ выяснилось, что это заболевание может быть прослежено в семьях пациентов на много поколений назад и что ХГ наследуется согласно определенному механизму: по крайней мере один из родителей каждого пациента страдал этим заболеванием и примерно половина детей этих больных также страдают им. Рис. 2.1 представляет собой иллюстрацию родословной семьи пробанда — носителя заболевания, страдающего ХГ.
ХГ передается по наследству как доминантный признак. Индивидуум, страдающий ХГ, является носителем одного доминантного аллеля (X), вызывающего развитие заболевания, и одного нормального (рецессивного) аллеля (х). Крайне редки случаи,
Рис,2.1, Пример родословной ее- когда пациент имеет два доминан-мьи,в которой хорея Гентингтона тных аллеля _ эта ситуация пред-передается по наследству (один из полагает, что оба родителя тако-родитеяей пробанда также страдает го пациента страдают ХГ. Люди, этим заболеванием). н£ страдающие хг обладают дву-
D -мужчина; о -Женщина; П-О- мя рецессивными аллелями (хх). супружеская пара; l-g3 - супружеская Родитель, страдающий ХГ, чаще пара и их ребенок; ♦ или ■-носитель всег0 является носителем геноти-заболсвания; ^Л или ^М — пробонд - Па Хх и в момент скрещивания носитель заболевания, через которого порождает гамету (яйцо или спер-были собраны сведения о родословной. мий\ либо с х, либо с х аллелем. (Подробнее о правилах составления гс- г
Фактических Древ см. гл. VII.) Гаметы нормального родителя
всегда содержат рецессивные аллели х. Четыре возможных комбинации этих аллелей показаны на рис. 2.2. Дети таких родителей всегда наследуют один здоровый аллель, передаваемый по наследству нормальным родителем. Однако, поскольку при зачатии аллели родителей комбинируются по случайному закону, для каждого из потомков вероятность наследования аллеля X от родителя, страдающего ХГ,
Рис.2.2. Схема скрещивания: аллельный механизм наследования хореи Гентингтона (пример доминантного наследования).
[Х] — доминантный аллель, вызывающий развитие ХГ; [х] — рецессивный аллель (здоровый).
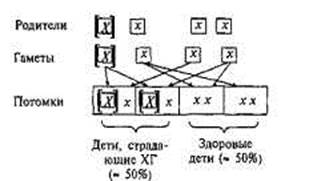
составляет 50%. Этим и объясняется тот факт, что у родителей, пораженных ХГ, только 50% детей страдают тем же заболеванием.
Для ХГ характерна одна особенность: первые симптомы этого заболевания проявляются лишь в зрелом возрасте, т.е. тогда, когда большинство людей уже создали семью и обзавелись детьми. В ином случае пациенты, страдающие ХГ, вообще не могли бы иметь потомков, так как умирали бы до наступления половой зрелости. Передача по наследству доминантного аллеля X возможна именно потому, что его летальный эффект не проявляется до начала репродуктивного периода.
Эта особенность развития ХГ создает чрезвычайно щепетильную психологическую ситуацию. В 1993 г. ученые открыли ген на хромосоме 4, вызывающий ХГ, и разработали молекулярно-генетический метод, позволяющий тестировать каждого человека с тем, чтобы определить, является ли данный индивидуум носителем патологического аллеля-мутанта (аллеля X).
Представьте себе следующую ситуацию. Ваши бабушка и дедушка по материнской линии умерли достаточно рано, и в семье не сохранилось никаких свидетельств того, что один из них, возможно, был носителем гена ХГ. Вашей матери 53, она больна ХГ. Вам 30, и у Вас есть возможность обратиться в лабораторию клинической генетики с тем, чтобы Вам сказали, являетесь Вы носителем гена X или нет. Вероятность того, что Вы — носитель этого гена, достаточно велика и составляет приблизительно 50%. Захотите ли Вы пройти подобный тест?
Исследования показывают, что большинство взрослых людей, для которых риск развития ХГ высок (поскольку один из родителей болен), предпочитают подобный тест не проходить. Этот тест, однако, имеет принципиально другое значение в пренатальной диагностике, когда заранее можно определить, является ли развивающийся организм носителем аллеля X. Ранняя пре-натальная диагностика позволяет родителям сделать осмысленный выбор относительно жизни их будущего ребенка, а также создает возможность раннего пренатального клинического вмешательства.
РЕЦЕССИВНОЕ НАСЛЕДОВАНИЕ: ФЕНИЛКЕТОНУРИЯ
Закон расщепления объясняет и наследование фенилкетонурии (ФКУ) — заболевания, развивающегося в результате избытка важной аминокислоты — фенилаланина (Phe) в организме человека. Избыток фенилаланина приводит к развитию умственной отсталости. Частота встречаемости ФКУ относительно низка (примерно 1 на 10 000 новорожденных), тем не менее около 1% умственно отсталых индивидуумов страдают ФКУ, составляя, таким образом, сравнительно большую группу пациентов, умственная отсталость которых объясняется однородным генетическим механизмом.
Как и в случае ХГ, исследователи изучали частоту встречаемости ФКУ в семьях пробандов. Оказалось, что пациенты, страдающие ФКУ, обычно имеют здоровых родителей. Кроме того, было замечено, что ФКУ чаще встречается в семьях, в которых родители являются кровными родственниками. Пример семьи пробанда, страдающего ФКУ,
оказан нарис. 2.3:больной ребенокродился уфенотипически здоровых родителей-кровныхродственников (двоюродных брата исестры), носестра отца ребенкастрадает ФКУ.
ФКУпередается порецессивному типу наследования, т.е.генотип больного содержит два аллеля ФКУ,полученные от обоихродителей. Потомки,
Рис. 2.3.Пример родословной семьи, вкоторой ФКУпередается по наследству (тетя пробанда страдает этим заболеванием).
Двойная линия между супругами обозначает кровнородственный брак. Остальные
обозначения те же, что и на рис. 2. 1
которые имеюттолько один такойаллель, нестрадают заболеванием, ноявляются носителями аллеля ФКУ и могутпередать егосвоим детям. Нарис. 2.4 показаны путинаследования аллелей ФКУ отдвух фенотипически нормальных
родителей. Каждый изродителей имеетодин аллель ФКУ иодин нормальный аллель. Вероятность того, чтокаждый ребенок можетунаследовать аллель ФКУ откаждого изродителей, составляет 50%.Вероятность того, что ребенокунаследует аллели ФКУ от обоихродителей одновременно, составляет 25% (0,5 х 0,5 = 0,25;вероятности умножаются, поскольку события наследования аллелей откаждого изродителей независимы друг отдруга). Ген ФКУ иего структурные варианты,встречающиеся в разныхпопуляциях, хорошо изучены. Знания,имеющиеся в нашемраспоря-
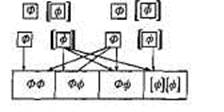
Родители
Гаметы
Потомки
Рис. 2.4. Схема скрещивания: аллельный механизм наследования ФКУ.
Ф — доминантный аллель («здоровый»); [ф]—рецессивный аллель, вызывающий развитие заболевания. ФФ, Фф — фенотипически нормальные дети (их 75%); только 25% имеют нормальный генотип (ФФ); еще 50% фенотипически здоровы, но являются носителями аллеля ФКУ (Фф). Оставшиеся 25% потомков — больны (1Ф11Ф1)-
жении, позволяют проводить своевременную пренатальную диагностику с тем, чтобы определить, унаследовал ли развивающийся зародыш две копии аллеля ФКУ от обоих родителей (факт такого наследования резко повышает вероятность заболевания). В некоторых странах, например в Италии, где частота встречаемости ФКУ достаточно высока, такая диагностика проводится в обязательном порядке для каждой беременной женщины.
Как уже отмечалось, ФКУ чаще встречается среди тех, кто вступает в брак с кровными родственниками. Несмотря на то что встречаемость ФКУ сравнительно низка, примерно 1 человек из 50 является носителем аллеля ФКУ. Вероятность того, что один носитель аллеля ФКУ вступит в брак с другим носителем такого аллеля, составляет примерно 2%. Однако при заключении брака между кровными родственниками (т.е. если супруги принадлежат к одной родословной, в которой аллель ФКУ передается по наследству) вероятность того, что оба супруга окажутся носителями аллеля ФКУ и одновременно передадут два аллеля будущему ребенку, станет значительно выше 2%.
Дата добавления: 2015-07-18; просмотров: 1647;