Особенности фенотипа ЭСК
В отличие от специализированных клеток ЭСК сохраняли уникальную потенцию наборами мРНК «ранних» генов эмбриогенеза. Редакторуя мРНК, ЭСК обходятся без многих блоков транссигнализации, которые используют дифференцированные клетки. ЭСК для контакта с микроокружением используют минимальное число рецепторов, сигнальных белков, транскриптаз хроматина . Между тем наборы " housekeeping genes", обслуживающих потоки энергии, веществ, метаболизм экспрессированы на уровне дифференцированных клеток. Второй важнейшей характеристикой незрелых ЭСК в культуре является высокий потенциал пролиферации. На поверхности пролиферирующих ЭСК экспонирован уникальный общий рецептор для LIF, SCF, IL-6 . Эта тройка сигналов через трансмембранную субъединицу GP-130 и соответственно Jak-Stat-3 транскрипционный комплекс процессирует сигналы в ядро, стимулируя вхождение G0 клеток в митоз. Схема событий между рецептором с ядром выглядят следующим образом. Высокоаффинное селективное связывание LIF, SCF, IL-6 с внешним рецептором активирует латентный фактор транскрипции Stat-3. Далее домен Stat-3 –SIE DNA binding –специфически взаимодействует с промотером c-fos на уровне ДНК хроматина (Рис 1-8). Примечательно, что активный Stat-3 не выявляли в эмбриобласте in situ. Активированный Stat-3 в комплексе с фактором тотипотентности Oct–4 запускал митозы и самообновление клонов ЭСК. Интенсивное самообновление ЭСК в культуре сопряжено с активацией тирозинфосфатазы SHP-2 Этот регуляторный фермент дефосфорилирует фосфотирозины разных белков (включая цитоплазматический сигнальный домен GP-130). Удаление фосфорных групп с тирозинов вызывает стойкую активацию проводящей субъединицы GP-130. Активация GP-130 стимулирует следующую киназу ERK (extracellular regulated kinase). SHP-2- зависимая активация ERK необходима в качестве кофактора для устойчивого самообновления ЭСК in vitro. Сохранение тотипотентности пролиферирующих мышиных линий ЭСК связано с присутствием в этих клетках дополнительного регуляторного белка Gab-1 - кофактора экспрессии главного белка тотипотентности Oct-4.
Похоже, что белок Oct4 выполняет разные функции в ранних зародышах млекопитающих. Избыток Oct4 в эмбриобласте и ЭСК блокирует развитие клеток трофобласта, поддерживая тотипотентность пролиферирующих незрелых клеток. С началом органогенеза Oct4 контролирует начало рестрикционного созревания многих клеточных линий. Oct4 необратимо исчезает в созревающих клетках. В неактивном виде Oct-4 выявлен о зрелых ооцитах, зиготе и эпибласте. В фетусе Oct-4 выявляется в половом бугорке и примордиальных половых клетках. Если в ЭСК увеличить экспрессию Oct-4, клетки спонтанно дифференцируются в эндодерму и мезодерму. Оказалось, что Oct-4 активирует одни, ингибируя другие гены в зародышевых тканях.
Бластоцисты мыши Oct4-/- останавливаются в развитии после имплантации. Такие аномальные зародыши преимущественно содержат клетки трофобласта, тогда как клетки эмбриобласта подвергаются гибели (Nicols J., Zevnik B., Anastassiadis K. Et al., 1998). Однако клетки трофобласта также вяло пролиферируют, поскольку в зародышах отсутствует FGF4 (индуктором экспрессии гена FGF4 является Oct4). Все зародыши-гетерозиготы Oct4-/+ проходят фазу имплантации без аномалий, поскольку 30-50% уровня Oct4 в эмбриобласте достаточно для его превращения в эпибласт и первичную полоску. Уровень Oct4 не влиял существенно на постимплантационную гибель зародышей.
Особую роль рецептор LIF/ GP-130 играет в задержке развития бластоцист у мышей, других грызунов на стадии диапаузы. Такие «законсервированные» на 3-4 нед бластоцисты, переживающие в полости матки, содержат зрелые клетки эпибласта вместо эмбриобласта. У зародышей с поврежденным GP-130 блокировано созревание эпибласта в диапаузных бластоцистах (Smith A.G., 2001)
Jak-Stat-3 "кабель" транссигнализации используется региональными гематогенными, мышечными стволовыми клетками для самообновления клеток клонов. LIF - рецептор ЭСК существенно влияет на выживание прогениторных клеток в культуре. Michel Revel из Вайссмановского института в Израиле показал, что прогениторные клетки в гематогенных клонах теряют способность формировать димерный вариант рецептора, который способен связывать IL-6. Такие клетки немедленно элиминируются из клона апоптозом. Если стволовые клетки трансфицировать генетической конструкцией, содержащей ген лиганда IL-6 и ген GP-130 рецептора, то трансфицированным клонам возвращается потенция к неограниченному самообновления с минимальным апоптозом. Такие клоны долгосрочно обновлялись in vitro и in vivo (Kollet O., Aviram R., Chebat J. et al., 1999).
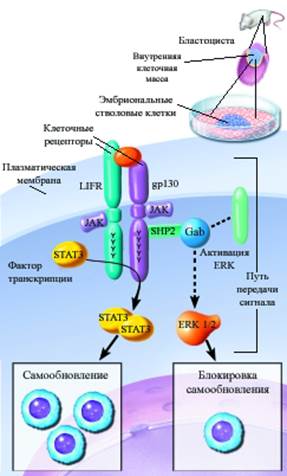
Рис 1-9. Система сигнализации LIF-рецептора в клеточное ядро
На следующем этапе прогениторные клетки пролиферуют под контролем спаренных рецепторов Delta/Notch в комплексе с фактором тотипотентности- продукта семейства генов Hes (Hes7 в клонах мезодермы, сомитах и клонах миогенных стволовых клеток скелетных мышц). В клонах стволовых клетках ЦНС экспрессирована пара Нes1, Hes5, тогда как один Hes5 экспрессирован в клетках- предшественицах олигодендроцитов.
"Минимальный фенотип" ЭСК и примордиальных половых клеток (ППК) проявлялся ошаренной формой клеток из-за отсутствия цитоскелета, белков и рецепторов адгезии на поверхности плазматической мембраны, а также многих кофакторов сигнализации, вмонтированных в компоненты цитоскелета. ЭСК и ППК имели высокую активность теломеразы и щелочной фосфатазы. В отличие от региональных стволовых клеток ППК не имели на поверхности бета1-интегрина. ППК вступают в дифференцировку, формируя вторичные агрегаты клеток в суспензии (эмбриоидные тельца).
В последние годы бессмертные тотипотентные линии ЭСК были выделены из эмбрионов крыс, коровы, приматов, свиней. Показательно, что все ЭСК имели не только близкое строение, но и практически универсальный набор антигенов-маркеров, (SSEA-3, SSEA-4, TRA-1-81, TRA-1-60). (Таблица 2)
| Таблица 2. Сравнительные характеристики фенотипа ЭСК разных видов | |||||
| Название маркёра | ЭСК/ППК/ТК Мыши | ЭСК приматов | ЭСК человека | ППК человека | ТК человека |
| SSEA-1 | + | - | - | + | - |
| SSEA-3 | - | + | + | + | + |
| SEA-4 | - | + | + | + | + |
| TRA-1-60 | - | + | + | + | + |
| TRA-1-81 | - | + | + | + | + |
| Щелочная фосфатаза | + | + | + | + | + |
| Oct-4 | + | + | + | Данные отсутствуют | + |
| Высокая активность теломеразы | + ЭСК, ТК | Данные отсутствуют | + | Данные отсутствуют | + |
| Зависимость от фидера | ЭСК, ППК, некоторые ТК | Имеется | Имеется | Имеется | Небольшая, низкая клоногенность |
| Факторы, способствующие самообновлению стволовых клеток | LIF и другие факторы, действующие через gp130- рецептор, способны замещать фидер | Сокультивирование с фидерными клетками; другие факторы не выявлены | Фидерные клетки + сыворотка; фидер + бессывороточная среда + bFGF | LIF, bFGF, форсколин | Факторы не выявлены, утеря тотипотентности в пассажах |
| Характеристики роста in vitro | Округлой формы, многослойные агрегаты ЭТ | Агрегаты ЭТ уплощённой формы | Агрегаты ЭТ уплощённой формы | Округлой формы, многослойные агрегаты ЭТ | Агрегаты ЭТ уплощённой формы |
| Формирование тератом in vivo | + | + | + | - | + |
| Образование химер | + | Данные отсутствуют | + | - | + |
Внешние домены этих "метчиков" представлены комплексом гликолипида GL7 с сиаловой кислотой. Фенотипические характеристики ЭСК и ППК не менялись после 20-30 пассажей. Культура продолжала расти плотными суспензионными клонами, в которых клетки удерживались плотными межклеточными контактами, резистентными к трипсину (рис 1-4 г-д).
Дата добавления: 2015-07-18; просмотров: 856;