Основные источники и способы выделения ЭСК
(историческая справка)
Хотя концепция стволовой клетки была предложена Александром Максимовым в 1908 г для кроветворной ткани, статус большой науки эта область получила в последнее десятилетие ХХ века. Первая попытка лабораторного оплодотворения яйцеклетки млекопитающих датирована 1878 г. Но лишь в 1959 г. в США был получен первый кролик путем искусственого оплодотворения. Первые природные тотипотентные клетки человека оказались в руках экспериментаторов только вначале 60-х годов.
В начале 70-х годов ХХ века Leroy Stevens обнаружил высокую частоту спонтанного возникновения тератокарцином в половых зачатках мышей линии 129. Эти опухоли для экономии он размножал в брюшной полости взрослых животных той же линии (а не в дорогостоящей культуре клеток). Среди конгломератов опухолевых клеток наблюдал появление неорганизованных популяций дифференцированных клеток: фрагментов кожи, хряща, волос, скопления миоцитов и кардиомиоцитов, кроветворной ткани (Репин В..С., 2000). Спонтанно возникшие тератокарциномы в культуре росли неприкрепленными клонами пролиферирующих плюрипотентных клеток, из которых часть подвергалась спонтанной дифференцировке в специализированные клетки (производные всех трех зародышевых листков). Стивенс первый высказал догадку, что дифференцированные клетки образуются не из раковых клеток, а из малой примеси пролиферирующих плюрипотентных половых зародышевых клеток, которые он описал как «эмбриональные стволовые клетки» (ЭСК) . С этой первой работы термин ЭСК прижился в литературе. В начале 70-х Stevens и Solter независимо друг от друга нашли второй источник эмбриональных стволовых клеток. Пассированную линию плюрипотентных эмбриональных клеток можно получить, если предимплантационные зародыши мыши вводить в брюшную полость, либо под кожу взрослым мышам. В 1975 году Минц доказала, что введение предимплнатационных зародышей мыши/крысы в любую ткань вне матки ведет к образованию опухоли из части клеток зародыша (эмбриокарцином). Высокоочищенные линии эмбриокарциномы (ЭК)были получены в результате многочисленных пассажей культуры через многие поколения животных. Авторы сразу обратили внимание на сходство поведения и фенотипа (ЭК) и тератокарцином (ТК). В культуре плюрипотентные клетки размножались клонами, причем часть клеток, покидавших клоны, подвергались разнообразной спонтанной дифференцировке. При этом клетки в клоне продолжали интенсивно самообновляться после многочисленных пассажей. Разными способами удавалось повышать число клон-инициирующих клеток в культуре. Таким образом было доказано, что плюрипотентность наследуется новыми поколениями клеток, возникающими в клоне, однако быстро теряется вне клонов. Упомянутые авторы первыми показали, что присутствие фидера (монослоя фетальных фибробластов) позволяет сохранять больше плюриптентных клеток в культуре. В дальнейшем из опухолей, размноженных пассажами через животных, удалось изолировать несколько линий эмбриокарцином. После Стивенса во многих лабораториях мира было изолировано более 100 линий ЭК (эмбриокарцином) и ТК (тератокарцином). Для лабораторных исследований линии мышиной тератокарциномы (129/sv, F19, F8, JM-1, E14TG2f, Zin40, CGR 86, R1, CCE) до сих пор остаются самой распространенной, дешевой моделью плюрипотентных клеток. Множество линий ТК и ЭК коммерциализованы биотехнологическими компаниями с готовым клеточным паспортом (иммунофенотип, хромосомный анализ, профиль экспрессии мРНК, профиль рецепторов и белков внутриклеточной сигнализации). Рядом авторов предложена классификация ТК как по происхождению, так и фенотипу клеток (Andrews P.W., Przyborski S.A., 2001). Существенный недостаток большинства линий ТК - быстрая утрата тотипотентности в пассажах, ограниченный потенциал цитодифференцировки клеток. Многие линии ТК оказались анеуплоидными, а потому генетически нестабильными (имеется в виду фентип клеток в пассажах), что снижает воспроизводимость результатов. Сопоставление фенотипа ЭСК и ТК привело Пирса к важной гипотезе о том, что многие опухоли возникают как ошибки созревания региональных стволовых клеток (Pierce, 1974). Для многих раковых линий характерна генетическая нестабильность, как и для анеуплоидных линий ТК. Вторым недостатком ТК и ЭКявляется риск малигнизации трансплантата. Третьим недостатком ТК и ЭК является смешанная дифференцировка прогениторных клеток в культуре. По этой причине линии ТК чаще всего использовались в опытах на животных в предклиническую фазу для изучения судьбы трансплантата (миграция, пролиферация, дифференцировка клеток, апоптоз, реваскуляризация трансплантата, сроки выживания в разных органах). Описано несколько линий ТКчеловека (N2, NTERA-1, NTERA –2). По соображения биоэтики не было попыток получения ЭКчеловека.. Однако высокоочищенные линии ТК человека, проверенные по стандартным критериям биобезопасности, уже используются в клинических испытаниях, в том числе в виде клеточных трансплантаций для коррекции клинических проявлений заболевания. Эти тщательно отобранные линии сохраняют стабильный кариотип, высокий потенциал цитодифференцировки и имеют высокий индекс встраивания в ранние зародыши..
В конце 70-х ХХ века Беатрис Минц и Карл Илменси из Ракового института (Филадельфия, США) первыми получили аллофенных зародышей мышей, смешивая в чашке петри клетки ТКс клетками нормальных предимплантационных зародышей (стадия 8-64 клеток). Зародыши - химеры из лабораторных и природных зародышевых клеток нормально развивались, проходили без аномалий внутриутробный период развития, а позднее нормально развивались в постнатальном периоде. Этим методом впервые получены межвидовые химерные организмы. Гетерогеномные зародыши реализовали один план морфогенеза без морфологических уродств. Пересадки ТКв предимплантационные зародыши не нарушали работу генов сегментации и гомеозиса, контролирующих 3D - карту зародыша. Это открытие сдалало возможным пересадки ТК для коррекции наследственных метаболических болезней, поскольку пересадки ЭСК не сопровождались аномалиями морфогенеза.
Использование фидерного слоя клеток для поддержания тотипотентности линий ТК оказалось полезным новшеством для следующего шага: выделения ЭСК из бластоцист мышей (Evans,Kaufman,1981; Martin,1981). Во-первых, клетки эмбриобласта нужно было отделить от трофобласта в культуре. Трофобласт мешал созреванию эмбриобласта в клетки эпибласта. Во-вторых, добавление 10-20% FCS+ меркаптоэтанол способствовало созреванию изолированных клеток эмбриобласта в клетки эпибласта. В-третьих, образование вторичных клонов эпибласта было необходимым условием возникновения ЭСК с неограниченным потенциалом пролиферации (Smith A.G., 2001).
Ранние (предимплантационные) зародыши и фетальная абортная ткань оставались главными природными источниками ЭСК. Зародыш на стадии бластоцисты представляет собой типичную «стволовую нишу», в которой впервые четко разделены тотипотентные стволовые клетки эмбриобласта и поддерживающие клетки трофобласта (своеобразный фидер) (Рис 1-5-А). Клетки трофобласта вырабатывали кофакторы выживания и защиты, необходимые для сохранения и пролиферации плюрипотентных клеток внутреннего зародышевого слоя. Одновременно они блокировали неконтролированную пролиферацию эмбриобласта (позже – эпибласта) in situ. Малая доля клеток эмбриобласта бластоцисты сохраняла тотипотентность (1-2% клеток удавалось переводить в бессмертную самовоспроизводящуюся линию ЭСК). Репрограммирование эмбриобласта в линию ЭСК начиналось с механического отделения эмбриобласта от трофобласта. Затем фрагменты эмбриобласта помещали на “фидер” из фетальных фибробластов без факторов, ограничивающих пролиферацию тотипотентных клеток. Первые попытки выделения ЭСК сделал Пирс в 1957 г. Историю вопроса можно найти в превосходном обзоре (AndrewsP.W., Przyborski S.A.,Thomson J.A., 2001).
В середине 70-х годов А.Я. Фриденштейн и сотр . показали, что строма гематогенной ткани взрослых мышей и человека содержит самообновляемые плюрипотентные клетки, которые можно клонировать в линии (Friedenstein A.J., Chailachyn R.K.,Latsinik N.V. et al., 1974, Friedenstein A.J.,Owen M.1988). В постмитотическом состоянии клетки этих линий дифференцировались в остеобласты, адипоциты, хондроциты, миоциты. Некоторые линии генерировали эндотелиоциты. В культуре эти линии формировали колонии фибробластоподобных клеток, хотя клетки размножались симметричными делениями (как прогениторные клетки). Фриденштейн назвал этот вид плюрипотентных клеток мезенхимальными стволовыми клетками. С этих пионерских работ началось изучение стволовых/прогениторных клеток мезенхимы.
Почему получение линий ЭСК мышей было сразу сфокусировано на клетках эмбриобласта? Во-первых, клетки этой ткани имели максимальное фенотипическое сходство с клетками ТК. Во-вторых, только дериваты этих тканей удавалось длительно пассировать в незрелом статусе, сохраняя после многочисленных пассажей способность дифференцировки в разные линии.. В третьих, исходные клетки эмбриобласта никогда не генерировали клеток трофобласта. Это разделение позволяло новым поколениям клеток избирательно встраиваться в морулу/бластоцисту и заселять ткани, возникающие из трех зародышевых листков. Если дериваты трофобласта начинали быстро генерировать полиплоидные клетки, то производные эпибласта автоматически поддерживали нормальный кариотип, были резистентны не только к аномалиям кариотипа, мутациям, но и многим эпигенетическим сигналам. Сразу после имплантации клетки эпибласта зародышей мыши подвергались интенсивной, но ограниченной пролиферации (Dani C., Chambers I., Johnstone S. et al., 1998).Если интенсивно пролиферирующие клоны эпибласта своевременно изолировали из зародыша, затем кондиционировали среду ростовыми факторами, то малая часть клеток приобретала способность к неограниченной пролиферации. Природа этой ключевой трансформации клеток эпибласта остается неразгаданной. Расшифровка сигналов, останавливающих пролиферацию клеток эпибласта в имплантированном зародыше, поможет эмпирическим поискам условий лабораторного и полу-производственного наращивания клеток эпибласта в автоматическом режиме. Уже очевидно, что ЭСК найдут широкое применение в качестве высокоаффиного вектора доставки новой ДНК в ранние зародыши как с исследовательскими, так и медицинскими целями.
Недавно было показано, что линии мышиных ЭСК с генотипом 40ХУ возникают легче, чем линии с генотипом 40ХХ. Более того, пересадки ЭСК с мужским генотипом 40ХУ в зону полового бугорка у зародыша самки вело к смене пола у развивающихся зародышей (Smith A.G., 2001).
В середине 80-х очередной методический прорыв осуществил Marius Capecchi, разработавший метод двойного выключения (нокаута) материнской/отцовской аллели гена в ЭСК мышей. Этот подход оказался незаменимым для изучения функции известных/неизвестных генов в раннем эмбриогенезе мышей. При смешивании 60-80 “нокаут”-ЭСК с 20-30 клетками нормальных ранних зародышей мыши получают развивающуюся химеру, закладки органов которой включают разные пропорции донорских/реципиентных клеток. С помощью таких химер открыты функции многих генов органогенеза, зародышевых листков, гомеозиса и сегментации. Метод двойного нокаута пока эффективно работает только на ЭСК мышей, но не других млекопитающих. За последние годы техника химеризации ранних зародышей продвинулась вперед с помощью трансфицированных донорских ЭСК, маркированных так называемыми генами цветных белков. Многие морские беспозвоночные используют язык флуоресцентных белковых молекул для коммуникации в абсолютной тьме на большой глубине океана. Донорские клетки, окрашенные разными белками, можно вводить в ранние зародыши и отслеживать судьбу живых клеток прямо под флуоресцентным микроскопом. Можно в ЭСК вводить лишний ген под бактериальным промотором, реагирующим на тетрациклин. Этим способом можно включать внесенный ген в зародышах, внося в среду антибиотик. Процесс химеризации зародышей можно прослеживать в динамике с момента введения донорских клеток, изучать пролиферацию, миграцию и судьбу донорских популяций в разных частях зародыша, в том числе в динамике методом цейтраферной съемки.
В настоящее время ЭСК мыши с двойным нокаутом практически любого гена поставляются на рынок биотехнологическими компаниями за 1500-2000 ам.долларов/ 2-3 млн клеток. За последние годы техника химеризации ранних зародышей донорскими стволовыми клетками достигла пределов эффективности. Так, пересадки одной гематогенной стволовой клетки человека в 8-клеточный зародыш мыши вызывали “гуманизацию” сразу нескольких органов мыши.
С помощью двойных делеций в ЭСК удалось получить мышей, копирующих мышечную дистрофию Дюшенна, наследственную атаксию-телангэктазию, другие наследственные фатальные заболевания детей. Современная биология пополнилась новым каталогом болезней мышей, копирующих основные наследственные болезни человека.
Мартин получил первую линию ЭСК мышей из бластоцисты в 1981 г. (Martin, 1981). В 1984-88 гг Эндрюс создал технологию получения "лабораторных" нейронов человека из линии тератокарциномы NTERA-2. В 1989 г описаны методы дифференцировки ЭСК тератокарциномы человека практически в любой тип специализированных клеток взрослого организма. В 1994 г Брижит Хоген из Вандербилтского ун-та, Нэшвилл, США опубликовала метод выделения примордиальных прогениторных клеток (ППК) из полового зачатка зародышей мыши. В 1995-96 гг Томсон изолировал бессмертные линии ЭСК из бластоцисты обезьяны.
Качественные сдвиги в этой проблеме возникли в 1998 г после изолирования бессмертных линий ЭСК человека. Решающую роль сыграла компания Герон из Калифорнии, которая вложила в исследования Томсона и Герхарда 5 млн долларов. Государственные академические учреждения наложили мораторий на эти исследования. Этот вклад фирмы обернулся новой биологией. Изолирование ЭСК человека авторитетный журнал Science назвал третьим по важности открытием в биологии ХХ века (после открытия двойной спирали и программы «Геном человека»). С середины 90-х годов не прекращались попытки получения линий ЭСК человека в нескольких лабораториях США, Великобритании, Канады, Индии, Австралии/Синапура, Японии.
В 1998 г ин-т репродуктивной биологии в Норфолке (Канада) первым наладил производство бластоцист человека из банка спермы и яйцеклеток. На втором этапе бластоцисты использовались для выделения линий ЭСК человека. Однако канадцы не успели первыми изолировать линию ЭСК человека из «лабораторных» бластоцист.
В 1998 г Джеймс Томсон (Висконсинский ун-т, США) изолировал 5 линий ЭСК из замороженных бластоцист человека, оставшихся неиспользованными после суперовуляции и получения оплодотворенных яйцеклеток с целью получения беременности (Thomson J., Itskovitz-Eldor J, Shapiro S.S., 1998). Оригинальный метод получения ЭСК из бластоцисты человека изложен в знаменитом патенте 6.200.806, полученном в марте 2001 г Wisconsin Alumni Res Foundation (WARF). Патент частично продан фирме Geron на получение некоторых специализированных клеток человека (нейроны, кардиомиоциты, клетки печени, поджелудочной железы).
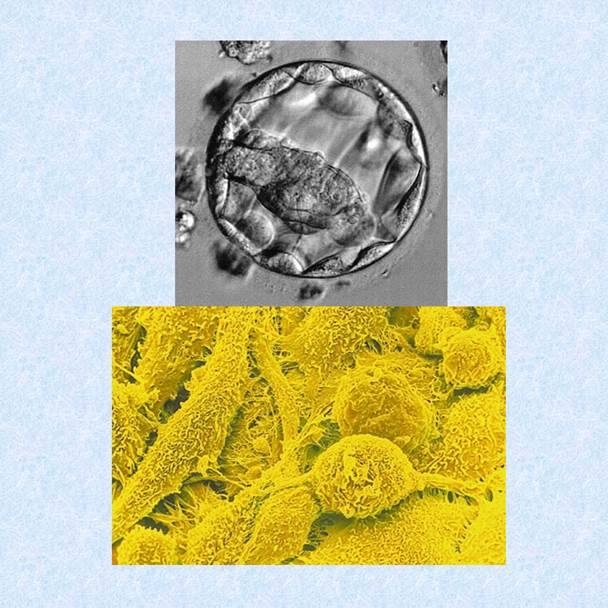
Рис 1-5 Устройство бластоцисты (А) - под ф/к микроскопом (Б) - ошаренные ППК на подложке из вытянутых клеток Сертоли (сканирующая электронная микроскопия)
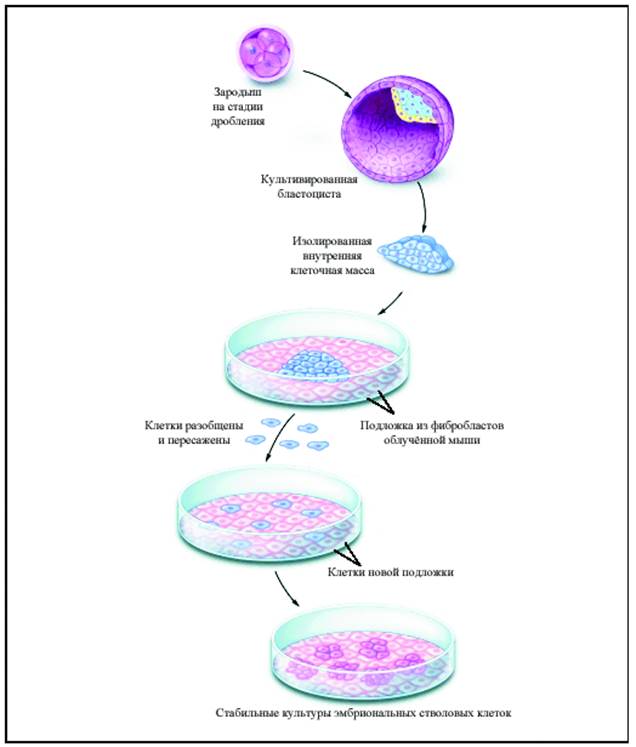
Рис 1-6. Выделение ЭСК из бластоцисты человека
Зародыши обязательно замораживали перед выделением ЭСК. Известно, что после замораживания большинство зародышей теряли способность проходить без отклонений послеимплантационный период развития в матке (одно из требований биоэтики для использования «остатков» зародышей после операции искусственного оплодотворения). После размораживания в среде Дульбекко с помощью игл и микроманипулятора выделяли эмбриобласт, предварительно помеченный флуоресцентно мечеными антителами. Мелко нарезанную ткань эмбриобласта выращивали в той же среде Дульбекко 9-15 дней над облученным фидером фетальных фибробластов с добавлением трех ростовых цитокинов (LIF, IL-6, SCF). Клетки в культуре интенсивно делились, формируя множество новых клонов по периферии микроэксплантатов эмбриобласта. Эти выросшие по краям эксплантата однородные колонии аккуратно собирали и повторно диспергировали (но не до одноклеточной суспензии). В суспензии ЭСК росли клонами, которые вновь диссоциировали пипетированием. Максимальную скорость экспансии клонов достигали повторной диссоциацией агрегатов на стадии 10-15 клеток. Новые клоны возникали через 5-7 суток. Каждый клон переносили в микроячейку и выращивали до агрегата из 40-50 клеток. Процедуру повторяли в пассажах, увеличивая плотность до 6-10 миллионов на чашку Петри (6 см). Из клонов удалось выделить несколько ХХ и ХУ линий ЭСК человека, которые через 100-120 пассажей сохраняли высокий темп клеточных делений, высокую активность теломеразы, минимальный белковый фенотип, тотальную потентность генома. Незрелые ощаренные клетки в мелких интенсивно пролиферирующих клонах экспрессировали на поверхности общий гетеродимерный рецепор для LIF, SCF и IL-6. В присутствии указанных митогенов сигнал от рецептора передается через трансмембранную субъединицу GP-130 в ядро. Детали устройства и работы этого рецептора показаны на рис (Рис 1-9). Активная сигнализация через рецептор в ядре блокировала остановку клеток в G0 -фазе, стимулируя немедленную инициацию G1 - фазы.
Дифференцировка ЭСК начиналась (после смены среды, удаления фидера и LIF, добавления сыворотки) с прикрепления клеток к подложке и формирования цитоскелета. Ни одна из линий не являлась клоном, т.к. была получена не из одной клетки (а из кластера клеток). Рост клонов шел периферией. В фазу равновесия пролиферация прогениторных слоев уравнивалась апоптозом и миграцией клеток из клона. Рост клоногенной культуры определяли тремя параметрами: а) числом клонов/мл, б) долей клеток в клоне/общее число клеток в культуре, в) скоростью обновления клонов в культуре. Линии ЭСК в культуре формировали суспензионные клоны двух типов: а) плотные симметричные сферы, б) рыхло собранные уплощенные колонии. Компактные сферы были резистентны к трипсину и пипетированию, тогда как рыхлые неправильные колонии диспергировали на отдельные клетки, как трипсином, так и механически.
В отличие от ЭСК мыши, линии ЭСК человека для сохранения пролиферации без дифференцировки требовали фидерного слоя клеток и LIF одновременно. Без LIF и фидера ЭСК прикреплялись к подложке, делились и формировали монослой из продвинутых диференцированных клеток. Сохранение потенциала пролиферации и незрелого фенотипа не являются частью автоматики этих клеток, а требуют специальных условий для реализации. При малых плотностях культивирования добавляли bFGF или сыворотку для поддержания пролиферации. Появился метод длительного пассирования (250 пассажей в течение года) линии ЭСК человека без фидера на поверхности матригеля в среде, кондиционированной супернатантом от выращивания фетальных фибробластов (Xu C., Inokuma M.S., Denham J. et al., 2001). Благодаря матригелю, удавалось нарастить больше клонов в единице объема. Поверхность микрочастиц матригеля была покрыта ламинином. ЭСК с помощью рецептора к ламинину прикреплялись к поверхности микрочастиц и формировали клоны (ламининовый рецептор – единственный рецептор клеточной адгезии, экспонированный на поверхности ЭСК человека и тератокарциномы). Даже при добавлении кондиционной среды после выращивания фетальных фибробластов и LIF, скорость роста клонов ЭСК оставалась невысокой (на порядок ниже, чем у Томсона). Без добавления супернатанта от фидера клоны ЭСК быстро теряли Oct-4 и вступали в скрытую фазу дифференцировки. Одиночные клетки после прикрепления автоматически дифференцировались. Образование смешанной культуры из дифференцированных распластанных по гелю клеток и клонов ЭСК было вторым недостатком метода.
Линии ЭСК человека, трансплантированные в виде суспензии одиночных незрелых клеток, генерировали тератомы в тканях взрослых иммунодефицитных животных (Odorico J.S., Kaufman D.S., Thomson J.A., 2001).
В 1998 г Джон Герхарт (Ун-т Джона Гопкинса, Балтимор, США) впервые изолировал бессмертные линии половых прогениторных клеток (ППК) из полового зачатка фетусов 4-5 недели гестации. Через 2-3 недели половой зачаток человека необратимо изменялся, хотя фенотипически сохранял те же клетки. Однако выделить ППК в виде бессмертной линии из зародышей более поздних сроков пока никому не удалось. ППК возникают в желточном мешке на 3-й неделе развития. Эти экстраэмбриональные провизорные клоны через 1-2 недели мигрировали в зону половых бугорков зародыша, где формировали "дормантные" популяции прогениторных плюрипотентных клеток. Значительная часть ППК сохраняется без изменений в зародыше вплоть до рождения. С ППК работать легче, поскольку этой ткани в фетусе больше, чем клеток эмбриобласта (Shamblott M.J., Axelman J., Gaerhart J. et al., 1998). Технология получения ППК запатентована в 1998 г (патент 6245566 от 12 июня 2001 г). Пересадки аллогенных ППК уже используют для лечения мужского бесплодия, поскольку донорские клоны ППК хорошо встраиваются и колонизируют иммунопривиллегированные вакантные зоны эпителия канальцев (Shinohara T., Avarbok M.R., Brinster R.L.,1999; Shinohara T., Orwig K., Brinster R.L., 2001). Данный метод особенно важен для получения зародышей-трансгенных химер, которые переносят новый генетический вектор в половые клетки и обеспечивают вертикальную передачу нового гена потомству. Существенно и то, что с фетальной абортной тканью можно работать в исследовательских лабораториях. Изолированную ткань полового зачатка 4-5 недельных фетусов диспергировали до взвеси одиночных клеток с помощью смеси коллагеназы IV-V, гиалуронидазы и ДНКазы. Ферментативное отделение ППК от стромальных клеток Сертоли необходимо для активации пролиферации ППК. Клетки Сертоли синтезируют набор антимитогенов и специальных факторов, удерживающих ППК в неактивном «дормантном» состоянии (Рис 1-6-Б)
Первичную культуру ППК также выращивали над слоем фидера (первый-второй пассаж фетальных фибробластов). Для стимуляции пролиферации ППК в среду добавляли bFGF, LIF и форсколин (стимулятор уровня цАМФ). В культуру ППК добавляли также 15% фетальную сыворотку (HyClone). Преобладающая часть клеток размножалась суспензионными клонами.
На следующем этапе изолировали самые крупные интенсивно пролиферирующие колонии с множественными пузырьками (кистами). Возникающие клоны являются истинными первичными клонами (в отличие от эмбриоидных телец, которые являются вторичными агрегатами клеток, вступающими в фазу дифференцировки ). Активно пролиферирующие клоны составляли примерно 2-5 % клеток от всей клеточной массы ППК. Лишь малый процент первичных клонов сохранял высокий темп пролиферации клеток. Эта спонтанно растущая плюрипотентная ткань содержала одновременно нестин+, виментин+, GFAP+ клетки (варианты нейрональных стволовых клеток). Эти же клеточные массы содержали стволовые гематогенные, мышечные, мезенхимальные стволовые клетки, маркеры предшественников эндотелия, а также маркеры эндодермы. Одновременно многие клетки в составе этих агрегатов имели экспрессированные мРНК трех зародышевых листков и мезензимы (Schamblott M.J., Axelman J.,Littlefield J.W., et al., 2001). Выращенная плюрипотентная ткань на стадии появления маркеров региональных стволовых клеток имела две особенности. Во-первых, пересадки этой ткани SCID- мышам не давали опухолей (в отличие от линий ЭСК). Во-вторых, ткань содержала мощные пролиферирующие клоны, которые сохраняли высокий темп делений через 50-70 пассажей. Исследования генетической тотипотентности эмбриоидных телец как in vitro, так и in vivo опровергли старую концепцию, что судьба стволовых клеток окончательно определена их положением в трех зародышевых листках или мезенхиме. Скорее высокая пластичность плюрипотентной ткани, возникающей первично из эмбриоидных телец, на основе синергизма стволовых клеток и сложных межклеточных взаимодействий - это пока предел возможного в биологии, установленный эволюцией. Именно этот суммарный потенциал стволовых клеток открывает новые возможности для регенеративной медицины.
Дифференцированные клетки, возникающие спонтанно при культивировании эмбриоидных телец (вторичных дифференцирующихся агрегатов), являлись главным блокатором выживания стволовых клеток. Своевременное удаление примеси дифференцированных клеток из эмбриоидных агрегатов восстанавливало нормальное обновление клонов и темп пролиферации (Mountford P., Nichols J., Zevnik B. et al., 1998).
Четвертый способ получения ЭСК человека был предложен сотрудниками Гарвардской медицинской школы в Армхерсте (Cibelli J., Stice I.C., Robl J.M. et al., 1998). С помощью электрического разряда соматические клетки фетуса (в ряде опытов- клетки самих исследователей) сливали с яйцеклеткой коровы, из которой был удален собственный пронуклеус. Такая "лабораторная химера", собранная из клеток двух видов млекопитающих, вела себя как зигота в культуре, развиваясь нормально до стадии бластоцисты (Kato Y.,Tani T., Sotomaru Y. et al., 1999).
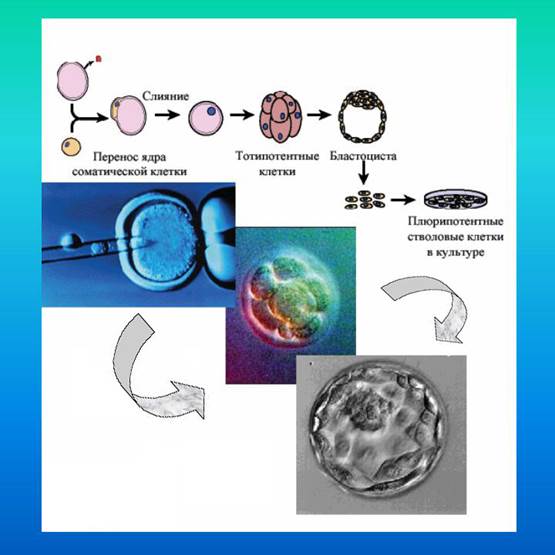
Рис 1-7. Получение ЭСК переносом ядра соматических клеток в оопласт
После имплантации в матку коровы такие "химерные зародыши" нормально проходили органогенез и заключительные фазы внутриутробного развития. В 1996 г студент-стажер Жозе Сибелли в лаборатории Джеймса Робла получил цитогибриды, собранные из ядра собственной соматической клетки, которое он пересадил в цитопласт коровы. Такие тотипотентные клетки в культуре стали развиваться до стадии бластоцисты. Результаты опытов были доложены президенту Клинтону, после чего на эти эксперименты был наложен мораторий.
Введение ЭСК крыс в зародыш мыши заканчивалось рождением мыши, органы которой были мозаично собраны из клеток крысы и мыши. Межвидовая химеризация зародышей млекопитающих породила множество нерешенных биоэтических и правовых вопросов. Потенции, генетические пути развития, возможности межвидовой сборки зародышей, особенно с использованием генома и ЭСК человека остаются неизученными. Второй способ клонирования зародышей связан с прямым переносом ядра соматической клетки (либо ядра ЭСК) в цитоплазму яйцеклетки того же вида, из которой был удален собственный пронуклеус. Вторая методика позволяла серийно клонировать бластоцисты (Wakayama T., Rodrigues I., Perry A.C. et al., 1999, 2001). Возраст донорских клеток существенно влияет на жизнеспособность и аномалии развития клонированных зародышей (Colman A., Kind A.,2000). Частота анеуплоидии, хромосомных аберраций и туморигенеза также возрастала в клонах с ДНК от клеток, взятых от пожилых доноров. Аномалии импринтинга в ранних зародышах, обусловленные нарушением гиперметилирования промотерных участков, служат ранним предвестником развивающейся генетической нестабильности клонов. Как известно, эффективность клонирования зародышей амфибий резко зависела от возраста клеток-доноров ДНК. Клонирование лягушек с помощью ядер эндодермы головастиков шло с эффективностью 2%, тогда как ядра клеток взрослых лягушек постоянно давали нулевой результат (Kikyo N., Wolffе A.P.,2000) . У мышей тотипотентность сохраняется до стадии 8 бластомеров. Только пересадки ядер зародышей до стадии 8-клеточных зародышей оказались эффективными для репродуктивного клонирования (полного воспроизводства зародыша и новорожденного по ДНК уже существовавших соматических клеток животного). Утеря тотипотентности резко снижала эффективность клонирования ранних и поздних зародышей вследствие множественных поломок эмбриогенеза. Поэтому методики репродуктивного клонирования целесообразно отлаживать и стандартизовать, используя максимально однородные ядра одной линии ЭСК (прежде всего мышей). До создания высокоэффективных и надежных методов репродуктивного клонирования преждевременно обсуждать проблему клонировании животных. Для проверки «репрограммирования» ядра соматической клетки до состояния тотипотентности предложено несколько методов, в том числе по способности рекапитулироватьь эмбриогенез мышиной бластоцисты после инъекции донорских клеток человека. Появление «гуманизированных» ростков клеток в зародыше и новорожденной мыши свидетельствует о тотипотентности генома созданных цитогибридов, полученных техникой пересадки ядра соамтической клетки (Cibelli J.B., Stice S.L., Golueke P.J. et al., 1998). Эффективность репрограммирования генома соматической клетки будет оцениваться независимо в промежуточном эксперименте, прежде чем выполнять весь эксперимент по репродукционноиу клонированию организма.
После переноса ядра ЭСК в яйцеклетку у части ядер достигалась максимальная тотипотентность к рекапитуляции начального и позднего эмбриогенеза, хотя в других тестах генетические потенции ядер ЭСК не были эквипотенциальны оплодотворенной яйцеклетке. ЭСК важны для клонирования редких животных. Вырожденность software органогенеза лучше изучать на предельно однородном генетическом материале (Kikyo N., Wolffe B., 2000).
Пересадки ядра дифференцированных клеток в цитоплазму ооцита сопровождаются потерей 75% ядерных белков (факторов транскрипции, белков-стабилизаторов гетерохроматина, тотальная смена гистонов-линкеров). Под влиянием нуклеоплазмина цитоплазмы ооцитов происходит удаление белкового каркаса ядер дифференцированных клеток, что ведет к деконденсации хроматина, дестабилизации нуклеосом и полной реорганизацией 3-D структуры хромосом. Импорт белков цитоплазмы ооцита автоматически ведет к реорганизации хроматина в состояние, близкое к тотипотентности, если используются ядра ранних зародышей. Однако полное автоматическое репрограммирование генома соматических клеток взрослых тканей оказалось пока технически невозможным (Kikyo N., Wolffe A.P., 2000)
Как известно, на лабораторное получение/разрушение зародышей человека для научных целей в США и ряде западных стран наложен мораторий. Линии тотипотентных клеток, дублирующие программы развития в обход беременности, вызывают в обществе много биоэтических сомнений, включая риск злоупотреблений. В США и ряде западных стран заморожен проект создания аутогенных ЭСК человека методом переноса ядра из соматической клетки. Кандидатами №1 на аутотрансплантацию признаны стволовые клетки взрослых пациентов, выделенные из кроветворной, жировой ткани, мезенхимы или волосяных фооликулов эпидермиса кожи. По мнению некоторых экспертов, мультипотентность генома этих клеток достаточна для наработки переживающих ростков донорской специализированной ткани. В 2001 г Vargaret Goodele и Karen Hirschi из Бэйлор колледжа (Хьюстон) показали, что трансплантация донорского костного мозга практически химеризует все органы реципиента. Донорские стволовые клетки в измеряемых количествах достигали поврежденного миокарда или поврежденной печени. В циркулирующей крови взрослых мышей, морских свинок, кролика были обнаружены клон-инициирующие МСК с остеогенным, хондрогенным, миогенным потенциалом. Однако таких клоногенных МСК практически не удалось обнаружить в циркулирующей крови у взрослых людей (Kuznetsov S.A., Mankani M., Gronthos S. et al.,2001). В работе британских исследователей удалось из крови нормальных взрослых людей выделить высокоочищенную фракцию CD34- клеток с варьирующим фенотипом и морфологией, которые удалось очистить на элютриаторе. Практически МСК составляют менее 1% очишенной фракции моноцитов. Однако МСК окрашивались антителами к виментину, CD-105 (эндоглину), коллагену I типа, рецептору витронектина и BMP-2 (Zvaifler N., Matinova L., Burger J. et al., 2000). В культуре пролиферирующие МСК формировали фибробластоподобные клоны. При добавлении индукторов дифференцировки клетки стромы формировали адипоциты, остеобласты или миофибробласты.
Первоначальный оптимизм в отношении возможности мезенхимы органов взрослых людей начал снижаться после открытия особенностей поведения МСК в тканях реципиента. Оказалось, что трансплантированные МСК сливаются с нейронами, миоцитами, гепатоцитами и другими соматическими дифференцированными клетками, формируя гетерокарионы. Такие гетерокарионы с маркерами МСК и фенотипом дифференцированных клеток принимали за трансдифференцированные МСК. Прежде всего в этой области необходима скрупулезная инвентаризация полиморфизма и функциональной гетерогенности так называемых резервных стволовых клеток мезенхимы, представленных множественными «нишами» в эпидермисе кожи, костно-мышечной, жировой, иммунной и кроветворной системе (Young H.E., Steele T.A., Bray R.A. et al., 2001) . Для определения доли стволовых/прогениторных МСК в разных тканях и органах взрослых организмов используют тест с инсулином и дексаметазоном. Все стволовые клетки в культуре под влиянием инсулина пролиферируют, не меняя незрелого фенотипа. Мишенью действия дексаметазона являются прогениторные клетки. Долю прогениторных клеток высчитывают по количеству возникших новых фенотипов клеток, опосредуемых эффектом стероида. Практически все пулы резервных МСК состоят из наборов стволовых/прогениторных субпопуляций с варьирующим фенотипом и генетической потентностью. Мультипотентность линий МСК сужается, особенно в пассажах. Первым исчезает хондрогенный потенциал линий МСК (Tsutsumi S., Shimazu A., Miyazaki K. et al., 2001) . Есть косвенные доказательства сохранения “дормантных” реликтов зародышевой мезодермы в мезенхиме взрослого организма, персистирующей in situ со стадии органогенеза (Young H.E., Duplaa C., Young M. et al., 2001; Zuk P.A., Zhu M., Mizuno H. et al., 2001). По фенотипу и плюрипотентности эти клетки невозможно отличить от МСК. Численность стволовых/прогениторных клеток в мезенхимальной ткани взрослого организма значительно больше, чем пул стволовых клеток в паренхиме (Deans R.J., Moseley A.B.,2000). Пока этому факту нет однозначного объяснения. Внутривенные вливания 50 млн МСК -дериватов гематогенной стромы, проведенные на здоровых реципиентах, признаны абсолютно безопасными (нет эффектов на клеточный состав крови, тромбоциты, иммунные клетки и иммунную систему) (Lazarus H.M., Haynesworth S.E., Gerson S.L. et al.,1995; Deans R.J., Moseley A.B., 2000).
Мультипотентность постнатальных стволовых клеток человека изучается на животных. Резервы стволовых клеток больных людей – прямой путь к банкам клеток, которые найдут более широкое применение, чем банки органов. Этот биоресурс незаменим в борьбе с фатальными заболеваниями органов. Даже в лидирующих странах система добровольного донорства качественных органов обеспечивает помощь 10-15% пациентам, находящимся на листе ожидания. Миллионы пациентов на планете погибают, не дождавшись кардинального лечения. С учетом нарастания эпидемии СПИДа, вирусного гепатита, новых форм особо опасныз вирусов, поражающих ниши стволовых клеток, растет потребность в альтернативных источниках клеток человеческих органов. Ткани взрослого человека становятся важным источником стволовых клеток для клиники. Микровкрапления эмбриональной зародышевой ткани сохраняются в костном, головном, спинном мозге, в криптах тонкого кишечника, волосяных фолликулах кожи, строме жировой ткани, в специальных стволовых пространствах глаза, периферических нервах, органах чувств. В здоровой ткани пул ЭСК обеспечивает медленную смену постаревших аномальных клеток на новые здоровые популяции. Как известно, апоптоз изношенных клеток в сочетании с обновлением клеточных популяций служит главным механизмом защиты от болезней (Wei G., Schubiger G., Harder F. 2000). Главная трудность в изучении СК взрослых органов – это преодоление их неактивного, «дормантного» состояния in vitro с целью наработки биотрансплантатов. Секреты макромасштабирования таких стволовых клеток – дело недалекого будущего. Получение GMP- трансплантов из ЭСК пациентов-заказчиков - новейшая страница бизнеса.
Пока мало известно, каким путем стволовые клетки нервной ткани контролируют обновление клеток ЦНС (McKey, 1997). Изолированные нейральные стволовые клетки (НСК) мышей линии Rosa, маркированные LacZ геном и бактериальной бета-галактозидазой, активно участвовали в образовании и заселении трех зародышевых листков и органов зародыша-реципиента. Пересаженные НСК частично мигрировали в костный мозг предварительно облученных животных, формируя колонии гематогенных клеток. Пересадки НСК в кровоток вели к образованию донорских колоний овальных (стволовых) клеток в дуктулярной системе печени. Существуют известные трудности в идентификации ЭСК in situ. Многие настаивают, что пролиферативный потенциал ЭСК взрослых тканей ограничен, и потому одновременно нужно создавать альтернативные источники СК для трансплантации.
Клоны стволовых мультипотентных клеток, выделенные из кожи взрослого человека, выделили в культуру с их последующей дифференцировкой в нейроны, глию, олигодендроциты (Toma J.G., Akhavan M., Fernandes K.J. et al., 2001). Стволовые клетки эпидермиса взрослых мышей линии С57ВL/6 извлекали из кожи и метили в культуре GFP (флуоресцентным белком). Далее меченые стволовые клетки кожи трансплантировали в изогенные бластоцисты. Затем химеры трансплантировали в матку гормонально подготовленных самок. Распределение донорских флуоресцентных клеток наблюдали на разных сроках пре- и постнательного развития.. Многочисленные ростки донорских клеток верифицировали в разных отделах головного мозга, нервном гребне, печени, почках, коже, других органах. Бластоциста мыши репрограммирует геном пересаженных региональных стволовых клеток взрослых тканей до статуса ЭСК (Liang L., Bickenbach J.R., 2002). Стволовые клетки из жировой ткани взрослого человека стали лабораторным источником дифференцированных линий многих органов и тканей человека (Zuk P.A., Zhu M., Huang J. et al., 2001). В перспективе жировая ткань и кожа взрослых пациентов окажутся важным источником стволовых клеток для аутогенной трансплантации.
| Таблица 1. Потенции и характеристики ЭСК взрослых тканей |

|
| Ограниченная пролиферация |
| Низкая теломераза |
| Мультипотентность генома |
| Полная иммуносовместимость |
| Создание банка клеток до собственного заболевания |
| Создание фамильного банка клеток |
|
|
Перспективным представляется пятый способ получения ЭСК из бластоцист-партеногенонов, полученных без спермиев и оплодотворения. Такие зародыши имеют диплоидный набор хромосом матери за счет сохранения второго пронуклеуса. Методы лабораторной активации яйцеклеток к «беспорочному» зачатию позволяют получать 20-30% бластоцист от общего числа яйцеклеток, активированых 1,2-пропандиолом и ионофором Са++ (Gook D.A., Osborn S.M., Johnston W.I., 1995; Rhoton-Vissak A., Lo P.Y., Barud K.M. et al., 1996). ЭСК, полученные из партено-бластоцист, сохраняют нормальную потенцию для крупномасштабной наработки специализированных соматических клеток.. Ткани-партеноты обнаружены случайной биопсией у взрослых людей без признаков болезни или функциональных нарушений (Surani M.A., 1995).
В 2001 г команда израильских и американских биологов расшифровала первые software для параллельного получения специализированных клеток экто-, мезо- и эндодермы из общего пула ЭСК мыши.(Schuldinger R., Yanuka O., Itskovitz-Eldor I. Et al., 2000). В 2001 г опубликован метод получения бета- и альфа-клеток островков Лангерганса из НСК головного мозга мышей (Lumelsky N., Blondel O., McKey R. et al, 2001). В конце 2001 объявлено о создании 60 бессмертных линий ЭСК человека (19 – в университете Гетеборга, Швеция, 9 - в компании CyThera, San Diego, California; 7 - Reliance Life Science, Mumbai, India, 6 - Monash University, Melbourn, Australia, 5 -Wisconsin Alumni Res. Foundation, 5 – Karolinska Universitst, Stockholm, 4 - Technion Israel Institute, Haifa, Israel ; 3 - National Center of Biological Studies, Bangalor,India; 2 – Unversity of California, San Francisco, USA). После проверок только 25 линий ЭСК отвечали выбранным критериям. Отобранные линии ЭСК имели сходную морфологию и молекулярные маркеры клеток, пассировались более одного года без изменения фенотипа м кариотипа клеток, без признаков бактериальной или вирусной контаминации. В августе 2001 г мультимиллионер Джим Кларк, руководитель Silicon Graphic, инвестировал 150 млн долларов в Центр стволовой клетки при Стенфордском ун-те для расшифровки главных программ, управляющих поведением ЭСК.
Дата добавления: 2015-07-18; просмотров: 812;