ПРИЧИНИ ВИНИКНЕННЯ СТОМЛЕННЯ
Що означає поняття стомлення під час фізичного навантаження? Відчуття стомлення під час виконання роботи до виснаження протягом 45-60 с, наприклад, забіг на дистанцію 400 м, дуже відрізняються від тих, котрих людина зазнає під час тривалого виснажливого м'язового зусилля, наприклад, марафонського бігу. Поняття стомлення використовується для характеристики загального відчуття стомленості, що супроводжується зниженням рівня фізичної активності.
Більшість спроб охарактеризувати й описати основні причини виникнення стомлення, а також місця її виникнення стосуються
• енергетичних систем (АТФ-КФ, гліколіз, окиснення);
• накопичення проміжних продуктів метаболізму;
• нервової системи;
• порушення скоротливого механізму волокон.
Жоден з цих параметрів окремо не може пояснити усі аспекти стомлення.
Наприклад, хоча відсутність необхідної кількості енергії може призвести до зниження здатності м'язів продукувати фізичні зусилля, енергетичні системи самі по собі не можуть бути причиною виникнення усіх форм стомлення. Так, відчуття стомлення, котрого ми часто зазнаємо наприкінці робочого дня, не має нічого спільного з наявністю АТФ. Стомлення може виникати і під дією стресу, довколишніх умов, що змінюють гомеостаз. На багато питань щодо стомлення допоки ще не отримано відповідей.
ЕНЕРГЕТИЧНІ СИСТЕМИ ТА СТОМЛЕННЯ
Енергетичні системи — це очевидна арена досліджень можливих причин виникнення стомлення. Коли ми відчуваємо стомленість, то часто говоримо: «У мене вичерпалася енергія». Однак таке застосування поняття «енергія» дуже далеке від його фізіологічного значення. Яку роль у вузькому розумінні відіграє енергія у виникненні стомлення під час фізичного навантаження?
Вичерпання запасів креатинфосфату. Згадаймо, що креатинфосфат використовується в анаеробних умовах для відновлення багатою енергією АТФ у міру її витрачення, отже, для підтримання запасів АТФ в організмі. Біопсичні дослідження м'язів стегна людини показали, що під час повторюваних максимальних скорочень м'язів виникнення стомлення співпадає з вичерпанням запасів КФ. Хоча такі види діяльності забезпечуються енергією АТФ, її запаси під час м'язових зусиль вичерпуються не так швидко, як запаси КФ, оскільки в утворенні АТФ беруть участь й інші системи. Однак після вичерпання запасів КФ здатність організму швидко поновлювати запаси використаної АТФ суттєво порушуються. Використання АТФ триває, але спроможність системи АТФ-КФ поновлювати використовувану АТФ значно зменшується. У результаті рівні АТФ також знижуються. У стані виснаження можуть бути вичерпані запаси як КФ, так і АТФ.
Щоб затримати виникнення стомлення, спортсмену необхідно контролювати інтенсивність зусиль, виходячи з найоптимальнішої швидкості виконання фізичної вправи, щоб не допустити передчасного вичерпання запасів КФ та АТФ. Якщо початкова швидкість дуже висока, то запаси КФ та АТФ швидко зменшаться, що призведе до ранньої стомлення та неспроможності зберігати швидкість на заключних стадіях дистанції. Тренування і досвід дозволяють спортсмену знаходити оптимальну швидкість, котра забезпечується найефективнішим використанням АТФ та КФ протягом усієї м'язової діяльності.
Вичерпання запасів глікогену. Підтримання рівнів АТФ забезпечується також аеробним та анаеробним розщепленням м'язового глікогену. У видах спорту, в котрих інтенсивні м'язові зусилля тривають усього декілька секунд, м'язовий глікоген є основним джерелом енергії для синтезу АТФ. На жаль, його запаси обмежені і швидко вичерпуються.
Як і у випадку з КФ, швидкість вичерпання запасів м'язового глікогену залежить від інтенсивності фізичного навантаження. Збільшення інтенсивності призводить до зниження рівня м'язового глікогену. Під час бігу на спринтерські дистанції, наприклад, м'язовий глікоген витрачається у 35-40 разів швидше, ніж при ходьбі. Наявність м'язового глікогену може бути обмежувальним чинником навіть за середніх зусиль. Для задоволення високої потреби м'яза в енергії під час фізичного навантаження необхідне постійне надходження глікогену.
Як видно з рис. 4.17, м'язовий глікоген швидше використовується у перші хвилини фізичної діяльності. Незважаючи нате що іспитований біг в одному темпі, інтенсивність використання глікогену у литковому м'язі була максимальною у перші 90 хв. Іспитований повідомляв про напруження, котрого він зазнавав (як важко йому було виконувати зусилля) у різні моменти тесту. Він зазнавав стресу середнього ступеня на самому початку, коли запаси глікогену були ще досить великими, незважаючи на високу інтенсивність його використання. Він не відчував значного стомлення, допоки запаси м'язового глікогену майже не вичерпалися. Таким чином, виникнення відчуття стомлення під час тривалого фізичного навантаження співпадає зі зниженням рівня м'язового глікогену. Марафонці називають раптове настання стомлення на 29-35-му кілометрі дистанції (18-22-га миля) «зіткненнямзі стіною». Часткововиникнення відчуття стомлення зумовлене вичерпанням запасів м'язового глікогену.
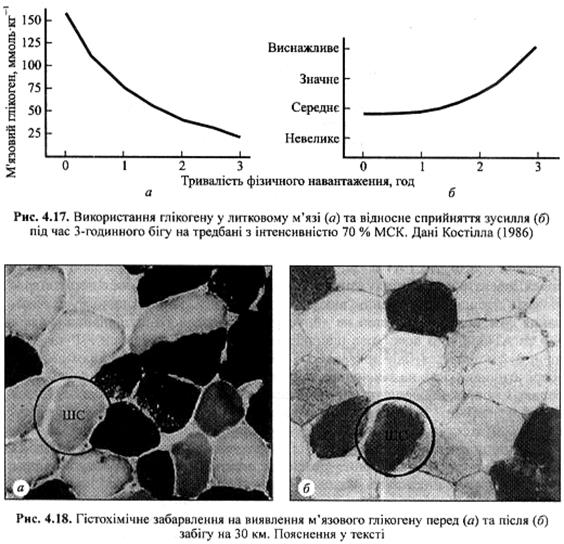
Вичерпання запасів глікогену у різних м'язових волокнах. Існують певні патерни рекрутування м'язових волокон та вичерпання у них запасів глікогену. Запаси глікогену можуть вичерпуватися у найчастіше використовуваних волокнах. Це призводить до скорочення числа волокон, здатних продукувати
 м'язове зусилля, необхідне для виконання фізичної вправи. На рис. 4.18 представлено мікроструктуру м'язових волокон, взятих з м'яза бігуна перед та після забігу на 30 км. На рис. 4.18, а ми бачимо диференціацію ПС та ШС волокон. Одне з ШС волокон обведене колом. На рис. 4.18, б продемонстровано другий зразок того самого м'яза, зафарбованого для виявлення глікогену. Чим червоніше (темніше) забарвлення, тим більше у ньому глікогену. До забігу усі волокна містили достатню кількість глікогену і мали червоне забарвлення. В обведеному колом ШС волокні на рис. 4.18, б (після забігу) все ще достатньо глікогену. Водночас у розташованих поряд з ним ПС волокнах запаси глікогену майже повністю вичерпані. Це свідчить про те, що ПС волокна більш інтенсивно використовуються під час м'язової діяльності, що потребує прояву витривалості, під час котрої продукується середнє зусилля, наприклад біг на 30 км.
м'язове зусилля, необхідне для виконання фізичної вправи. На рис. 4.18 представлено мікроструктуру м'язових волокон, взятих з м'яза бігуна перед та після забігу на 30 км. На рис. 4.18, а ми бачимо диференціацію ПС та ШС волокон. Одне з ШС волокон обведене колом. На рис. 4.18, б продемонстровано другий зразок того самого м'яза, зафарбованого для виявлення глікогену. Чим червоніше (темніше) забарвлення, тим більше у ньому глікогену. До забігу усі волокна містили достатню кількість глікогену і мали червоне забарвлення. В обведеному колом ШС волокні на рис. 4.18, б (після забігу) все ще достатньо глікогену. Водночас у розташованих поряд з ним ПС волокнах запаси глікогену майже повністю вичерпані. Це свідчить про те, що ПС волокна більш інтенсивно використовуються під час м'язової діяльності, що потребує прояву витривалості, під час котрої продукується середнє зусилля, наприклад біг на 30 км.
Патерн вичерпання запасів глікогену у ПС та ШС волокнах залежить від інтенсивності фізичного навантаження. Згадаймо, що ПС волокна першими залучаються при невисокій інтенсивності фізичного навантаження. Зі збільшенням інтенсивності починають включатися і ШСа волокна. Коли інтенсивність зростає до максимальної, то настає черга ШСб волокон. Вірогідно, що такий самий патерн є характерним і для процесу вичерпання запасу глікогену.
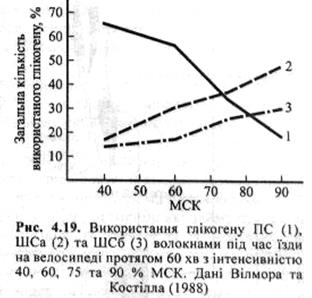
На рис. 4.19 показано кількість глікогену, котрий використали ПС, а також ШСа та ШСб волокна латерального широкого м'яза стегна під час їзди на велосипеді з різним відсотком МСК іспитованого. При відносно невисокій інтенсивності фізичного навантаження (40-60 % МСК) найактивнішими були ПС волокна. При вищій інтенсивності (75-90 % МСК) частіше використовувалися ШС волокна, котрі швидше вичерпували свої запаси глікогену порівняно з ПС волокнами.
Це, однак, не означає, що ПС волокна менше використовуються під час максимальних скорочень, ніж ШС, просто тут відображено той факт, що ШС волокна більшою мірою «покладаються» на глікоген. При інтенсивних м'язових скороченнях у роботі беруть участь усі типи волокон. Коли у ПС волокнах вичерпуються запаси глікогену, то ШС волокна стають або нездатними продукувати достатнє зусилля, або не можуть бути повністю рекрутованими, щоб компенсувати недостатнє зусилля. Тому висувалося припущення, що відчуття м'язового стомлення або важкості при тривалому фізичному навантаженні можуть відображати нездатність деяких м'язових волокон реагувати на вимоги, що ставляться фізичним навантаженням.
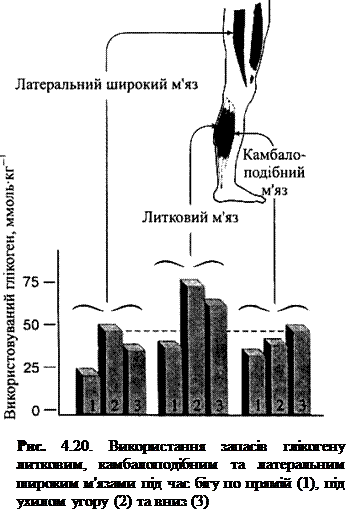 Вичерпання запасів глікогену у різних м'язових групах. Окрім вибіркового вичерпання запасів глікогену у ПС та ШС волокнах, фізичні навантаження можуть ставити надвисокі вимоги до окремих м'язових груп. Розглянемо дослідження, у котрому іспитовані бігали на тредбані, встановленому у трьох положеннях — горизонтально, з нахилом униз і з нахилом угору, — протягом 2 год при 70 % МСК. Як показують результати (рис. 4.20), незалежно від того, чи біжить спортсмен по похилій угору, униз чи горизонтально, найбільше глікогену використовують литкові м'язи. Це свідчить про те, що м'язи-розгиначі гомілковостопного суглоба, що піддавалися найбільшому фізичному стресу під час бігу на довгі дистанції, найскоріше вичерпують запаси глікогену на відміну од м'язів стегон. Таким чином, ділянкою виникнення стомлення єм'язи нижньої частини ніг.
Вичерпання запасів глікогену у різних м'язових групах. Окрім вибіркового вичерпання запасів глікогену у ПС та ШС волокнах, фізичні навантаження можуть ставити надвисокі вимоги до окремих м'язових груп. Розглянемо дослідження, у котрому іспитовані бігали на тредбані, встановленому у трьох положеннях — горизонтально, з нахилом униз і з нахилом угору, — протягом 2 год при 70 % МСК. Як показують результати (рис. 4.20), незалежно від того, чи біжить спортсмен по похилій угору, униз чи горизонтально, найбільше глікогену використовують литкові м'язи. Це свідчить про те, що м'язи-розгиначі гомілковостопного суглоба, що піддавалися найбільшому фізичному стресу під час бігу на довгі дистанції, найскоріше вичерпують запаси глікогену на відміну од м'язів стегон. Таким чином, ділянкою виникнення стомлення єм'язи нижньої частини ніг.
Вичерпання запасів глікогену та глюкоза крові. Один лише м'язовий глікоген нездатний забезпечити достатньої кількості вуглеводівпри фізичних навантаженнях, щотривають кілька годин. Значну кількість енергії під час фізичних навантажень, котрі потребують виявленнявитривалості, забезпечує глюкоза,що надходить у м'язи з кров'ю. Запаси глікогену в печінці розщеплюються, що забезпечує постійне надходження глюкози у кров. У першімоменти виконання вправи для утворення енергії необхідно відноснонебагато глюкози крові, однак пізніше внесок глюкози в утворенняенергії значно збільшується. Щобзабезпечити адекватне споживанняглюкози м'язами, із збільшеннямтривалості фізичного навантаженняпечінці доводиться розщеплюватиусе більше й більше глікогену.
Запаси глікогену у печінці обмежені, і вона не може швидко розщеплювати глюкозу з інших субстанцій. Отже, коли споживання м'язами глюкози перевищує утворення її печінкою, рівні глюкози крові знижуються. Не маючи можливості отримати достатню кількість глюкози з крові, м'язи змушені більш інтенсивно використовувати свої резерви глікогену, що прискорює вичерпанняйого запасів у м'язах і призводить до раннього виникнення стомлення.
Вплив глікогену на м'язову діяльність. Не дивує той факт, що збільшення постачання м'язів глікогеном на початку фізичного навантаження на витривалість поліпшує результати. Значення накопичення м'язового глікогену для м'язової діяльності, що потребує виявлення витривалості, обговорюється у розділі 14. Поки відмітимо, що виснаження запасів глікогену та гіпоглікемія (знижений вміст глюкози у крові) обмежують рівень м'язової діяльності у видах спорту аеробної продуктивності. Стомлення у видах спорту коротшої тривалості, мабуть, є результатом акумуляції проміжних продуктів метаболізму,
таких, як лактат та Н+, у м'язах.
ПРОМІЖНІ ПРОДУКТИ МЕТАБОЛІЗМУ ТА СТОМЛЕННЯ
Згадаймо, що молочна кислота — це проміжний продукт гліколізу. Хоча багато хто вважає, що саме вона є головною причиною стомлення та виснаження при усіх видах фізичних навантажень. Молочна кислота акумулюється у м'язовому волокні тільки під час короткочасного м'язового зусилля високої інтенсивності. У марафонців, наприклад, рівні молочної кислоти наприкінці забігу можуть бути майже такими, як у стані спокою, незважаючи на крайній ступінь стомлення. Як уже відмічалося, виникнення стомлення у марафонців зумовлене недостатнім енергетичним забезпеченням, а не надлишком молочної кислоти.
У спринтерів (бігунів, плавців, велосипедистів) під час змагань спостерігається швидка акумуляція молочної кислоти. Однак наявність молочної кислоти сама по собі не є причиною стомлення. Якщо вона не виводиться, то розщеплюється, перетворюючись на лактат, і веде до акумуляції іонів водню, що, у свою чергу, призводить до підкиснення м'язів і насамкінець — до ацидозу. Результати у видах спорту короткої тривалості та високої інтенсивності м'язової діяльності, такі, як біг і плавання на спринтерські дистанції, багато у чому залежать від інтенсивності гліколізу. У спортсменів, які займаються цими видами спорту, утворюється велика кількість лактату та Н+ у м'язах. На щастя, клітини та рідини організму мають буферні системи, такі, як бікарбонат (НСО3−), котрі зводять до мінімуму негативний вплив Н+. Якщо б не було таких буферів, то Н+ призвів би до зниження рН до 1,5 та порушення життєдіяльності клітини. Завдяки буферним системам організму концентрації Н+ залишаються невисокими навіть під час виснажливих фізичних навантажень. Тому м'язовий рН знижується від показника, характерного для стану спокою — 7,1 до 6,6-6,4 у стані виснаження.
Однак подібні змінення рН негативно впливають на утворення енергії та м'язові скорочення. Внутрішньоклітинний рН нижчий за 6,9 гальмує дію фосфофруктокінази — важливого глюколітичного фермента, що сповільнює інтенсивність гліколізу та утворення АТФ. При рН 6,4 вплив Н+ припиняє подальше розщеплення глікогену, викликаючи різке зниження рівня АТФ і насамкінець — стомлення. Окрім того, Н+ може витісняти кальцій з волокон, втручаючись у процес сполучення поперечних містків актину та міозину та знижуючи скоротливу силу м'язів. Багато учених вважають, що низький м'язовий рН є головним чинником, котрий лімітує м'язову діяльність, а також основною причиною виникнення стомлення під час короткочасних максимальних фізичних навантажень.
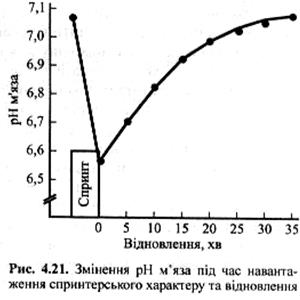 Як видно з рис. 4.21, відновлення після виснажливого фізичного навантаження спринтерського характеру рівня м'язового рН відбувається протягом 30-35 хв. Навіть при відновленому рівні рН концентрації лактату у крові та м'язах можуть залишатися підвищеними. Однак спортсмен може продовжувати виконувати вправу з відносно високою інтенсивністю навіть при показникові м'язового рН нижче за 7,0 та рівні лактату крові вище за 6-7 ммоль∙л -1, тобто такому, що у 4-5 разів перевищує рівень у стані спокою. Відновлення, хв.
Як видно з рис. 4.21, відновлення після виснажливого фізичного навантаження спринтерського характеру рівня м'язового рН відбувається протягом 30-35 хв. Навіть при відновленому рівні рН концентрації лактату у крові та м'язах можуть залишатися підвищеними. Однак спортсмен може продовжувати виконувати вправу з відносно високою інтенсивністю навіть при показникові м'язового рН нижче за 7,0 та рівні лактату крові вище за 6-7 ммоль∙л -1, тобто такому, що у 4-5 разів перевищує рівень у стані спокою. Відновлення, хв.
Останнім часом деякі тренерита спортивні фізіологи намагаються на основі вимірювання показників лактату крові визначити інтенсивність та обсяг тренувальних занять, що забезпечують оптимальні тренувальні стимули. Такі вимірювання дозволяють визначити інтенсивність тренувань, одна вони навряд чи зможуть охарактеризувати анаеробні процеси або ступінь ацидозу м'язів. Оскільки лактат та Н+ утворюються у м'язах, дифундуючи з клітин у позаклітинні рідини організму, потім транспортуючись до інших ділянок тіла для наступного обміну, то показники лактату крові залежать від інтенсивності його утворення, дифузії та окиснення. На ці процеси може впливати безліч чинників, тому доцільність використання показників лактату для визначення обсягу та інтенсивності тренувальних занять є дещо спірною.
НЕРВОВО-М'ЯЗОВЕ СТОМЛЕННЯ
До цього часу ми розглядали у межах м'язів чинники, котрі можуть бути причиною виникнення стомлення. Однак ряд даних вказує, що за певних обставин стомлення може виникати внаслідок нездатності активувати м'язові волокна. Це є функцією нервової системи. Як розглядалося, нервовий імпульс передається по кінцевій пластинці рухового нерва до м'яза, щоб активувати м'язову мембрану та змусити саркоплазматичний ретикулум виділяти кальцій. У свою чергу, кальцій, з'єднуючись із тропоніном, ініціює м'язове скорочення. Обговоримо два можливих нервових механізми, котрі можуть порушити цей процес та сприяти виникненню стомлення.
Нервова передача. Стомлення може виникнути на кінцевій пластинці рухового нерва м'яза, запобігаючи передачі нервового імпульсу до мембрани м'язового волокна. У дослідженнях, що проводилися на початку XX століття, спостерігали подібну нездатність передачі нервового імпульсу у стомленому м'язі. Вона може бути пов'язана з одним або кількома такими процесами:
• виділенням або синтезом ацетилхоліну — нейромедіатора, що переключає нервовий імпульс з рухового нерва на мембрану м'яза, кількість котрого може бути знижена;
• холінестераза — фермент, що розщеплює ацетилхолін після того, як той переключив імпульс, може стати гіперактивною, запобігаючи утворенню достатньої концентрації ацетилхоліну для ініціації потенціалу дії;
• активність холінестерази може знизитися (гіпоактивність), внаслідок чого відбудеться надмірне накопичення ацетилхоліну, що паралізує волокно;
• може підвищитися поріг збудження мембрани м'язового волокна;
• деякі субстанції можуть стати на боротьбу з ацетилхоліном за рецептори на мембрані, не активуючи її;
• калій може покинути внутрішньоклітинний простір скоротного м'яза, знижуючи наполовину потенціал мембрани.
Більшість із цих причин нервово-м'язової блокади пов'язана з нервово-м'язовими розладами (наприклад, міастенією), але вони так само можуть зумовлювати деякі форми нервово-м'язового стомлення. Існує також припущення, що стомлення може виникати у результаті затримки кальцію у Т-трубочках, що призводить до зменшення кількості кальцію, необхідного для здійснення м'язового скорочення. У дійсності вичерпання запасів КФ та накопичення лактату можуть просто збільшити інтенсивність акумуляції кальцію у Т-трубочках. Ці теорії виникнення стомлення поки що залишаються лише припущеннями.
Центральна нервова система (ЦНС) також може бути місцем виникнення стомлення. Є дані, що підтверджують, і дані, що заперечують це. Перші дослідження показали, що коли м'язи іспитованого опиняються на межі крайнього стомлення, то підтримка словом, окрик або пряма електростимуляція м'язів можуть збільшити силу м'язових скорочень. Ці дані свідчать про те, що лімітуючими чинниками м'язової діяльності при виконанні виснажливого фізичного навантаження можуть бути і психологічні чинники. Механізми, що лежать в основі стомлення ЦНС, недостатньо вивчені. Неясно також, чи обмежена ця форма стомленості тільки ЦНС, чи пов'язана з периферичною передачею нервових імпульсів.
Залучення до роботи м'язів частково залежить від свідомого управління. Психологічна травма виснажливого фізичного навантаження може свідомо або несвідомо пригнітити схильність спортсмена зносити біль. ЦНС може знизити інтенсивність навантаження до стерпного рівня, щоб «захистити» спортсмена. Учені єдині у тому, що дискомфорт стомлення, котрого зазнають, передує виникненню фізіологічних обмежень у м'язах. Більшість спортсменів, якщо тільки для них не є характерним високий рівень мотивації, припиняють виконання вправи, перш ніж настане фізіологічне виснаження їхніх м'язів. Щоб досягти піка м'язової діяльності, спортсмени змушені на тренуваннях відпрацьовувати виконання вправ у найоптимальнішому темпі, а також тренувати толерантність до стомлення.
Дата добавления: 2015-08-26; просмотров: 2584;