Особенности функциональной анатомии поперечнополосатых и гладких мышц
Скелетные (поперечнополосатые) мышцы состоят из множества функциональных единиц – мышечных волокон (клеток). Они имеют цилиндрическую форму и тесно прилегают друг к другу. Диаметр клеток поперечнополосатых мышц составляет от 0,005 до 0,1 мм, а длина может достигать нескольких сантиметров. Пучки мышечных волокон окружены соединительнотканными прослойками. На конце мышц коллагеновые волокна соединительной ткани образует сухожилия, которые служат для прикрепления мышц к разным частям скелета.
Общее число соединительной ткани в скелетной мышце довольно значительно. В зависимости от типа мышцы содержание коллагена в ней колеблется от 30% до 3% (рис. 37).
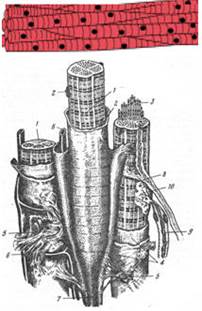
Рис. 37. Схема строения поперечно-полосатой мышечной ткани.
1 - мышечные волокна; 2 - ядра; 3 - миофибриллы; 4 - сарколемма; 5 - эндомизий; 6 - кровеносные капилляры; 7 - сухожильная нить; 8 - вегетативное нервное волокно; 9 - соматическое (двигательное) нервное волокно; 10 - аксоно-мышечный синапс [78]
Соединительнотканные прослойки мышцы обильно иннервируются рецепторами (колбами Краузе, тельцами Фатера-Пачини, ноцицепторами). Они располагаются и в фасциальных нервных стволах, и в жировых дольках.
Поперечнополосатые мышцы вместе со связками и периартикулярными тканями составляют около половины массы человеческого тела.
Мышечная клетка (волокно) одета прозрачной оболочкой – сарколеммой. Последняя по структуре подобна мембране нервной клетки и играет важную роль в возникновении и проведении возбуждения.
Цитоплазма мышечной клетки (саркоплазма) состоит из саркоплазматического матрикса – однородной среды, в которую погружены миофибриллы – сократимая субстанция неживой природы. Одно волокно содержит до 1000 фибрилл [68].
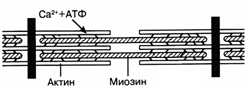
Рис. 24.Схема сокращения актин-миозинового комплекса
Каждая из миофибрилл состоит из саркомеров, образованных белковыми нитями двух типов - актином и миозином. При сокращении мышцы укорачиваются лишь волокна миозина. Они разбухают в поперечном направлении благодаря поглощению воды. При растяжении мышцы миозиновые столбики удлиняются. Сокращение актин-миозинового комплекса возможно только при присоединении ионов C2+ к особому регуляторному белку тропонину, локализованному на миозиновых нитях. Кроме того, для этого необходимо расщепление одной молекулы АТФ (рис. 38).
Мышечные волокна, осуществляющие непосредственное сокращение мышцы, называются экстрафузальными. Различают ещё и специализированные – интрафузальные - мышечные волокна, которые непосредственно в сокращении мышцы не участвуют. Они входят в состав сенсорных образований мышцы или её проприорецепторов – мышечных веретён.
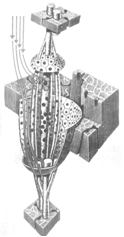
Рис. 25. Модель типичного мышечного веретена, сокращение саркомера [255]
В каждой мышце насчитывается от 90 до 100 мышечных веретён, которые расположены параллельно экстрафузальным мышечным волокнам и являются своего рода датчиками растяжения мышцы. Типичное мышечное веретено состоит из нескольких интрафузальных волокон, заключённых внутри толстой соединительнотканной капсулы (рис. 39).
Экстрафузальные мышечные волокна получают двигательную иннервацию от альфа-, а интрафузальные – от гамма-мотонейронов переднего рога спинного мозга. Кроме того, интрафузальные волокна веретён снабжены ещё и специализированными чувствительными (афферентными) нервными окончаниями.
Суть рефлекса на растяжение мышцы (миотатического рефлекса) заключается в следующем: любое увеличение длины мышцы сопровождается растяжением веретён. При этом происходит возбуждение афферентов. Возникающий импульс приходит к альфа-мотонейронам, вызывая сокращение всей мышцы за счёт её экстрафузальных волокон. Степень сокращения определяется величиной предшествующего растяжения.
Вследствие действия внешних факторов (инерция, гравитация) миотатический рефлекс минимальной степени выраженности наблюдается постоянно во всех скелетных мышцах, даже в условиях полного покоя и играет важную роль в поддержании и регуляции мышечного тонуса [60].
Помимо сократимости, важными биомеханическими свойствами скелетных мышц являются упругость и растяжимость.
Эластические структуры мышцы аккумулируют энергию упругих деформаций и частично возвращают её во время движения. Например, в ахилловом сухожилии может накапливаться потенциальной энергии упругой деформации больше, чем в икроножной мышце. Однако длительно растянутая и затем отпущенная мышца возвращается к своей исходной длине не полностью, а остается некоторое время более или менее удлиненной. Все эти явления связаны с вязкоупругими свойствами соединительнотканных элементов мышцы [33].
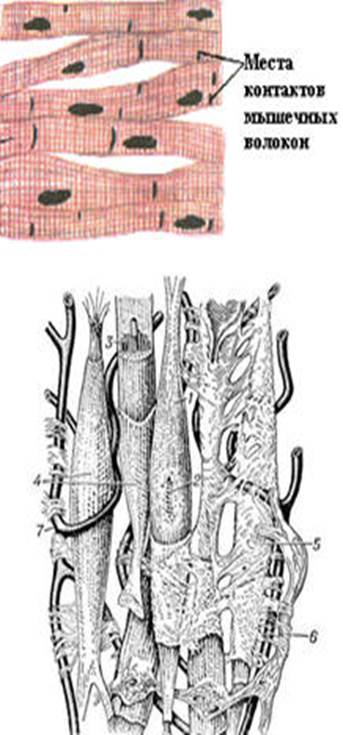
Рис. 26. Схема строения гладкой мышечной ткани.
1 - гладкая мышечная клетка; 2 - её ядро; 3 - миофибриллы; 4 - сарколемма; 5 - соединительная ткань; 6 - нерв; 7 - кровеносный капилляр [78]
Гладкая мышечная ткань, так же, как и поперечнополосатые мышцы, имеет клеточное строение. Её клетки толщиной 2-10 мкм имеют веретенообразную форму и небольшую, по сравнению со скелетными волокнами, длину – 50 - 400 мкм.
Гладкие мышцы контактируют друг с другом отростками так, что мембраны двух клеток соприкасаются. Зоны контакта занимают 5% поверхности клеточной мембраны. Здесь происходит передача возбуждения от одной клетки к другой. Между гладкомышечными клетками вплетены волокна соединительной ткани, образующей плотный футляр (рис. 40) [191].
Советскими учёными А. А. Заварзиным, Н. Г. Хлопиным и др. установлено, что гладкие мышцы – группа различных по происхождению тканей, объединяемых единым функциональным признаком – способностью к сокращению. У позвоночных гладкие мышцы слюнных, потовых и молочных желёз происходят из эктодермы, а внутренних органов – из мезенхимы [79].
Гладкомышечные волокна обладают высокой чувствительностью к различным БАВ: адреналину, норадреналину, ацетилхолину, гистамину и др., а также к таким факторам, как РО2, РС02 и Ph. Это обусловлено наличием специфических рецепторов на их мембранах.
Сократительный аппарат гладких мышц не обладает столь упорядоченным строением, как в поперечнополосатых мышцах. Миофибриллы состоят из нитей актина и миозина, которые идут в различных направлениях и прикрепляющихся к разным участкам сарколеммы.
Укорачиваются гладкие мышцы также за счет взаимодействия актиновых и миозиновых нитей друг с другом. Однако расщепление АТФ, необходимое для этого процесса, происходит в 100 - 1000 раз медленнее, чем у поперечнополосатых мышц. Поэтому гладкие мышцы способны долго находиться в состоянии сокращения, затрачивая сравнительно мало энергии и не подвергаясь утомлению.
В ответ на растяжение гладкая мышца сокращается. Это вызвано тем, что растяжение изменяет мембранный потенциал клеток, увеличивает частоту потенциалов действия и в конечном итоге повышает её тонус. Кроме того, гладкомышечные клетки сами являются рецепторами растяжения.
В организме человека это свойство служит одним из способов регуляции двигательной деятельности внутренних органов. Например, увеличение тонуса желудка в ответ на растяжение способствует сохранению объема органа и лучшему контакту его стенок с поступившей пищей. В кровеносных сосудах растяжение, создаваемое колебаниями кровяного давления, является основным фактором миогенной саморегуляции тонуса сосудов.
Еще одной специфической характеристикой гладкой мышцы является вязкость. Если её длительно удерживать в состоянии удлинения, то напряжение будет постепенно падать [240].
Таким образом, по своим основным свойствам – способности к сократимости (спонтанной или инициированной растяжением) и вязкости гладкие мышцы принципиально не отличаются от поперечнополосатых.
Дата добавления: 2015-07-14; просмотров: 2787;