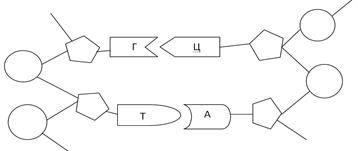
Схема двойной спирали ДНК
|
| |
| |
| |
| |
|
| |
| |
| |
| |
-Аденин
-Гуанин
-Тимин
-Цитозин
При раскручивании спирали водородные связи между азотистым основаниями разрываются и две цепочки ДНК расходятся. На каждой цепи из структурных элементов ДНК – дезоксирибонуклеозитрифосфатов – синтезируются новая /дочерняя/ цепь; при этом с каждым из нуклеиновых оснований спаривается комплементарное ему основание, так что каждая из двух новых цепей вместо одной двойной спирали появляются две точные репликации ДНК гарантирует сохранение генетической информации.
Транскрипция ДНК. Возникает еще вопрос: каким образом содержащаяся в генах информация преобразуется в аминокислотную последовательность белка, как определяется такая информация активность и другие свойства ферментов?
ДНК будучи носителем наследственной информации , тем не менее сама не служит матрицей для синтеза полипептидов.
Биосинтез белков происходит на рибосомах, которые непосредственно с ДНК не соприкасаются. Передачу, записанной в ДНК информации к местам синтеза белка осуществляет матричная, или информационная рибонуклеиновая кислота /М-РНК/.
РНК – полимер, отличающейся от ДНК лишь заменой дезоксирибозы на рибозу, а тимина /Т/ на урацил /У/. По строению очень напоминает одноцепочную ДНК. Матричная РНК синтезируется на одной их цепей ДНК, причем механизм этого процесса сходен с механизмом репликации ДНК. По последовательности нуклеотидных оснований ее цепь комплементарна цепи ДНК. Таким образом, при синтезе м-РНК просто копируется нуклеотидная последовательность ДНК. Этот процесс называют транскрипцией и противопоставляют его трансляции – переводу нуклеотидной последовательности в последовательность аминокислот.






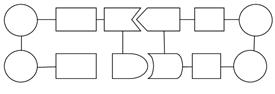
 Транскрипция м-РНК Трансляция Белок
Транскрипция м-РНК Трансляция Белок
Репликация ДНК
Обратная Транскрипция
Генетический код. Каждый ген представлен определенным участком молекулы ДНК. Специфическая информация, содержащая в гене, определяется последовательностью оснований в цепи молекулы ДНК.
«Алфавит», с помощью которого записана эта информация ДНК, включает четыре «буквы» - основания аденин /А/, гуанин /Г/, тимин /Т/ и цитозин /Ц/. В м-РНК тимин заменен урацилом /У/.
Специфичность ферментных белков синтез которых контролируют гены, определяется последовательностью аминокислот в полипептидных цепях. Эта же последовательность определяет и пространственную структуру белка, т.н. конформацию /вторичную, третичную и четвертичную структуру/.
Для перевода с языка нуклеиновых кислот на язык аминокислот служит специфический код. Каждая аминокислота определяется группой из трех соседних нуклеотидов – триплетом, или кодоном.
Та или иная последовательность триплетом в НК однозначно определяет последовательность аминокислот в полипептидной цепи. В триплетах возможно 64 различных комбинаций нуклеотидов. Если бы каждая из 20 аминокислот кодировалась лишь одним единственным триплетом, то 44 комбинации остались бы неиспользованными.
Но оказалось, что многие аминокислоты кодируются двумя и большим числом различных триплетов. Некоторые триплеты имеют особый смысл - они означают «начало» или «конец» полипептидной цепи. Триплеты считывают как 1,2,3; 1,2,3 и т.д. от начала молекулы м – РНК.
Трансляция м – РНК и синтез белка. Аминокислоты соединяются в полипептидную цепь в порядке, определяемой триплетами м – РНК. В этом процессе участвуют и РНК, транспортные РНК /т-РНК/, ряд ферментов, АТФ и другие факторы. Активация и присоединение аминокислот к соответствующей т-РНК осуществляется с помощью специфического фермента – аминоацил – м РНК – синтетазы, которая распознаёт, с одной стороны аминокислоту, а с другой соответствующую т-РНК. Имеется 20 различных синтетаз. Рибосома перемещается вдоль м РНК и таким образом происходит наращивание очередных аминокислот и формирование полипептидной цепи.
Одновременно происходит закручивание полипептидной цепи и свертывание её в клубок, определение также последовательностью аминокислот и природой их боковых цепей /гидрофобные и гидрофильные группы/. В результате возникает структура, обуславливающая свойства и функцию данного белка.
К м РНК обычно прикрепляется несколько рибосом и на одной и той же матрице одновременно синтезируется несколько полипептидных цепей. Такой комплекс одной м РНК с рибосомами называют полисомой. На конце полипептидной цепи от рибосомы.
Таким образом, нуклеотидная последовательность ДНК представляет собой закодированную «инструкцию», определяющую /при посредстве м РНК/ структуру специфического белка. Представление о передаче информации от ДНК через РНК на белок называют «центральной догмой» молекулярной биологии.
Таким путем происходит перенос информации у всех организмов, у которых генетическим материалом служит ДНК. Этот универсальный процесс передачи информации при репликации ДНК, транскрипции и трансляции представлен на приведенной выше схеме. Эта схема применима к эукариотам, прокариотам и ДНК-вирусам.
Но среди вирусов есть такие, у которых РНК реплицируется прямо на матрице РНК. Однако, у некоторых онкогенных РНК-вирусов вначале происходит синтез ДНК, контролируемый РНК, т.е. РНК служит матрицей для синтеза ДНК.
Такая информация, содержащаяся в вирусной РНК передается на ДНК путем обратной транскрипции /при помощи фермента обратной транскриптазы/. Он находит применение в генной инженерии. У прокариотических организмов обратная транскрипция не найдена.
Процесс транскрипции находится под строгим контролем, в результате количество молекул м РНК в клетке, коплементарных разным генам сильно различается. Хотя механизмы синтеза ДНК и РНК сходны, процесс транскрипции не обладает той степенью точности, которая характерна для репликации ДНК.
Поскольку м РНК не способна к самовоспроизведению, возникающие при ее синтезе ошибки в последующих клеточных генерациях не воспроизводятся и, следовательно, не могут наследоваться.
Механизм трансляции также отличается высокой точностью, но вероятность ошибки в целом выше, чем в случае синтеза ДНК и РНК. Наиболее уязвимый этап – “узнавание” с помощью фермента аминокослоты соответствующей молекулы т-РНК. Частота возникновения ошибок на этом этапе составляет 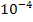 , что и определяет возможно, уровень точности процесса синтеза белка в целом. Но эти ошибки не воспроизводятся, если она не закодированы исходно в генетическом материале – ДНК.
, что и определяет возможно, уровень точности процесса синтеза белка в целом. Но эти ошибки не воспроизводятся, если она не закодированы исходно в генетическом материале – ДНК.
Таким образом, процессы транзикции и трансляции, служащие для выражения в онтогенезе генетической информации, не приводят к наследованию изменений, возникающих при их функционировании.
Только изменения, происходящие в молекуле ДНК, могут сохраняться в ряду поколений, поскольку они воспроизводятся в процессе репликации.
Следовательно, в основе эволюции прокариот, появлении новых видов, подвидов, штаммов и вариантов лежит способность к изменению только их генетического материала. У прокариот весь генетический материал, необходимый для жизнедеятельности, представлен одной хромосомой т.н. бактериальная клетка гаплоида. Но в определенных условиях /высокий уровень метаболизма и роста/ в клетках бактерий может содержаться по нескольку копий хромомсомы, что необходимо учитывать в селекционной работе.
Кроме того, у многих бактерий обнаружены внехромомсомные генетические элементы – плазмиды. Это кольцевые ковалентно замкнутые молекулы ДНК, содержащие от I500 до 40,0 тыс. пар нуклеотидов, реплицирующихся автономно как единое целое. К настоящему времени плазмиды обнаружены у I35 видов, принадлежащих к более чем 40 родам бактерий.
Обычно о присутствии плазмид в бактериальной клетке судят по проявлению определенных новых признаков, которые присущи этим структурам, т. е. кодируются их генетическим материалом. К таким признакам относится устойчивость к лекарственным препаратам, способность к переносу генов при конъюгации, синтез веществ антибактериальной природы, способность использовать некоторые сахара или обеспечивать деградацию ряда веществ.
Дата добавления: 2015-07-14; просмотров: 1708;