Время, мин
__VО2 .-.-. КрФ
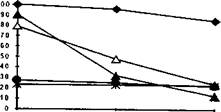
Р'ис. 4. Динамика возрастания потребления кислорода (V02) и скорости КФК-реакции в зависимости от предельного времени упражнения (по /Волков И.И., 1964])
Однако, как уже отмечалось выше, анализ большого числа проведенных отечественных и зарубежных исследований не позволил обнаружить ни одной работы, в которой путем прямых измерений или путем моделирования было бы показано, что: 1) парциальное напряжение кислорода внутри мышечных волокон (рО2) в начальный период работы любой мощности; 2) рО2 при работе максимальной аэробной мощности; 3) рО, при работе выше максимальной аэробной мощности у тренированных спортсменов падает ниже критического для митохондрий уровня (0,1-1 мм рт. ст. [Физиология человека / Под ред. Р. Шмидта и Г.Тевса. -Т.З., 1996]) или, что поток кислорода в митохондрии в состоянии их максимальной активизации [Gnaiger T. и др., 1997] ограничивает скорость выработки энергии при парциальном напряжении О2 в 1-3 мм рт.ст., которое, как можно предположить, наиболее типично для максимально активированной мышцы. Это же можно сказать и для степени насыщения миоглобина кислородом, высокое значение которой свидетельствует о доступности кислорода для протекания окислительно-восстановительных реакций в митохондриях. Так как миоглобин является депо кислорода внутри мышечного волокна, осуществляет «облегченную диффузию кислорода» [Холлоши Дж.О., 1982] и степень насыщения которого не должна снижаться, видимо, ниже 17-19%, что наблюдается в некоторых участках сокращающегося волокна при максимальном потреблении кислорода этим волокном и соответствует напряжению кислорода 1 мм рт. ст. [Gayeski T.E.J., C.R.Honig 1986].
Хотя, разумеется, в связи с ускорением потребления кислорода митохондриями его тканевое напряжение падает ниже уровня покоя, что является совершенно необходимым приспособительным механизмом, который обуславливает увеличенный градиент по кислороду между саркоплазмой МВ и капиллярной кровью, т.е. ускоренную доставку кислорода внутрь МВ при учете роли оксимиоглобина в качестве депо и переносчика О2 [Физиология человека/ Под ред. Р. Шмидта и Г.Тевса. -Т.З., 1996; Gayeski T.E.J., C.R.Honig, 1986]. Следовательно, есть основания говорить о наличии т.н. функциональной гипоксии, однако нет никаких прямых или косвенных данных, свидетельствующих, что степень такой тканевой гипоксии оказывается на- столько высокой, что ограничивает скорость выработки
энергии в митохондриях. Это справедливо (при нормальном парциальном напряжении 02 в атмосфере), по меньшей мере, и тех случаях, когда:
—нет нарушений в системе микроциркуляции, как при
сосудистых заболеваниях;
—нет функциональной недостаточности сердечной мышцы, вызванной, например, относительно небольшим (менее 3-3,5 см2) «устьем» аорты [47];
- нет состояния полной детренированности сердечной мышцы, которое может наблюдаться в экспериментах с гипокинезией или после продолжительной болезни;
— отсутствует ишемическая гипоксия, связанная с длительным напряжением мышц или короткими паузами расслабления мышц.
Кроме того, гипотеза об «инерционности» ССС не согласуется с тем, что при резком психоэмоциональном стрессе или в начале мышечной работы предельной интенсивности [Волков II.И., 1969], все параметры деятельности ССС возрастают по гораздо более крутому градиенту, чем в рассматриваемом примере. Представления об «инерционности» ССС также просто противоречат здравому смыслу — зачем организму в начале работы создавать себе трудности, накапливая «дефицит» кислорода, а затем «напрягаться» для того, чтобы его ликвидировать. Тем более, что существует механизм (см. ниже) который вполне позволяет обойтись без этих проблем. Правда, чтобы правильно интерпретировать сказанное, надо ясно понимать разницу между «энергозапросом», т.е. запросом в АТФ и «запросом кислорода», т.е. в количестве кислорода, которое способны использовать митохондрии при данном уровне стимуляции тканевого дыхания и который, естественно, различается в разных мышечных волокнах в зависимости от их активности, от их метаболического профиля, в зависимости от стадии развертывания внутриклеточных реакций энергообеспечения и т.д. Нам кажется, что эти два понятия постоянно перепутываются.
В подтверждение высказанной мысли, что в период врабатывания мышечные волокна не испытывают нехватки кислорода для полноценного тканевого дыхания из-за «инерционности» ССС, можно привести следующие расчеты.
При максимальной мощности работы мышц (например, в спринтерском беге или тесте на велоэргометре) снижение на-
блюдаемой мощности начинается на 5-6 с работы (рис. 4), однако, приблизительно до 8-10-й с человек еще способен поддерживать практически максимальную мощность, после чего начинается ее быстрое снижение [Волков Н.И., 1969; Nevill М.Е. и др., 1996]. Проведенные исследования со взятием биопсийных проб [Nevill М.Е. и др., 1996] показали, что к этому моменту концентрация КрФ снижается лишь на 50 - 55% (в мышце с 50% ММВ). В более «быстрой» мышце, вероятно, может быть обнаружена более низкая концентрация КрФ — до 75% [Greenhalf P.L.и др., 1987]), что заведомо выше значений, фиксируемых в конце более продолжительной работы [Greenhalf P.L. и др., 1996; Nevill М.Е. и др., 1996] и выше критической величины (в среднем по мышце), при которой энергии фосфагенов начинает не хватать для обеспечения максимальной мощности и силы взаимодействия миофиламентов [Greenhalf P.L. и др., 1996] . Для того чтобы снять это противоречие примем во внимание следующие четыре факта:
1) скорость расходования КрФ пропорциональна АТФ-аз-
ной активности миозина или креатинфосфокиназы [Хочачка
П., Дж. Семеро, 1988; Hirvonen J. и др., 1987; Greenhalf P.L. и др., 1996], которые максимальны у быстрых гликолитических мышечных волокон (БгМВ);
2) концентрация КрФ в целом пропорциональна массе со-
кратительных белков мышц (актина), хотя и может несколько
увеличиваться при тренировке [Биохимия: Учебник для инсти-
тутов физ. культуры, 1986; Яковлев Н.Н., 1983];
3) относительная сила быстрых и медленных МВ в мышце,
или не различается [Биохимия: Учебник для институтов физ.культуры, 1986], или на 10-30% выше у БМВ. С учетом п. 2, это означает, что запасы КрФ в быстрых и медленных МВ различаются в предельном случае не более чем на 40% (в среднем 10-30%) [Greenhalf P.L. и др., 1996; Nevill М.Е. и др., 1996];
4) соотношение АТФ-азной активности миозина у ММВ и
БгМВ определено как 1 : 3-4 [Биохимия: Учебник для инсти
тутов физ. культуры, 1986] (реально скорость расхода КрФ в ММВ медленнее, чем в БМВ только в 2-3 раза и это соотношение зависит от времени после начала максимальной работы [Greenhalf P.L. и др., 1996; Nevill М.Е. и др., 1996]).
Из приведенных данных следует очень существенный для предмета нашего исследования вывод — время снижения кон-
центрации КрФ до критического уровня зависит от АТФ-азной активности миозина и активности КФК, поэтому если при максимальной мощности волокон запасов этого субстрата в БгМ В хватает на 5-6 с [Hirvonen J. и др., 1987] (хотя реально скорость КФК реакции начинает снижаться уже со второй с максимальных тетанических сокращений [Greenhalf P.L. и др., 1996], то в ММВ его запасов должно хватать на 15-20 с. Этот вывод в целом подтверждается известной схемой Н.И. Волкова [1964] о динамике вклада КФК-реакции в энергообеспечение мышечной работы и данными электростимуляции мышц (когда расход субстрата идет с максимальной скоростью во всех типах МВ) (рис. 5) [Greenhalf P.L. и др., 1996].
|
| Изометр. сипа |

|
Однако при интерпретации данных рис. 5 следует учитывать различную длительность активных и пассивных фаз в реальных локомоциях и при электростимуляции. Другими словами, начало существенного снижения максимальной мощности около 8-12-й с работы связано с исчерпанием КрФ главным образом в БгМВ, в то время как в ММВ, у которых активность АТФ-азы ниже, а запасов КрФ не намного ниже (на 10-30% - см. выше), концентрации КрФ должно вполне хватать для обеспечения достаточно высокой мощности работы этих волокон в течении 15-20 с, даже без привлечения дополнительных источников АТФ, которыми являются гликолиз и окислительное фосфорилирование.
| О с. |
| 20 с. |
Дата добавления: 2015-08-21; просмотров: 913;