ТИПЫ СОХРАННОСТИ ИСКОПАЕМЫХ
Объектами палеонтологических исследований являются любые ископаемые биологического происхождения. К ним относятся не только сами организмы, но и биогеохимические компоненты, возникшие в биосферах прошлого при участии организмов. Для всех них широко используют термины «органические остатки», «ископаемые», «окаменелости» и «фоссилии» (лат. fossilis — погребенный, ископаемый). Последний термин в настоящее время очень популярен в зарубежной и русскоязычной литературе, хотя его использовали уже во времена Екатерины II.
Фоссилизация. Процессы преобразования погибших организмов в ископаемые называют фоссилизацией. Гибель и последующее захоронение организмов сопровождается воздействием различных факторов среды. Они проходят все процессы диагенеза, т. е. физических и химических преобразований при переходе осадка в породу, в которую они заключены.
После гибели организма в первую очередь разрушаются мягкие ткани, затем начинается заполнение пустот скелета вмещающим осадком или минеральными соединениями. Иногда пустоты скелета подвергаются пиритизации, ожелезнению, часто в них возникают друзы и щетки кальцита, аметиста, флюорита, галенита и т. д. Ископаемые скелеты нередко оказываются заключенными в фосфоритовые конкреции. При фоссилизации скелеты подвергаются перекристаллизации, приводящей к более устойчивым минеральным модификациям. Например, арагонитовые раковинки преобразуются в кальцитовые. Нередки случаи минерализации, когда первичный химический состав скелета изменяется (псевдоморфозы). Так, известковые раковины частично или полностью замещаются водным кремнеземом и наоборот. Также наблюдаются фосфатизация, пиритизация и ожелезнение минеральных и органических скелетов.
Растения при фоссилизации обычно разрушаются полностью, чему способствуют процессы гниения и брожения. Тем не менее ископаемые растения обнаружены начиная с докембрия. Чаще всего от растений сохраняются обугленные остатки листьев, лис-топодобных образований, стеблей, стволов, корней, семян, плодов, шишек, спор и пыльцы. В процессе фоссилизации первичные растительные ткани могут разрушаться полностью, и тогда остаются отпечатки и ядра. Нередко при фоссилизации растительные ткани замещаются различными минеральными соединениями, чаще всего кремнеземом, карбонатом и пиритом. Подобное полное или частичное замещение стволов растений при сохранении внутренней структуры называется петрификация. Чаще всего окаменение стволов связано с замещением кремнеземом, реже пиритом, кальцитом и др. Известны скопления стволов, так называемые окаменелые леса. Это либо целые стволы, либо только их нижние части.
Некоторые органические образования растений (воск, смола, лигнин, целлюлоза) сохраняются в ископаемом состоянии, почти не изменяясь. Минеральные слабоизмененные компоненты растений встречаются также довольно часто: это кремневые раковинки диатомовых водорослей, известковые «плодовые шарики» харовых растений, известковые пленки и желваки красных водорослей и т. д.
В зависимости от полноты сохранности и своеобразия остатков выделяют следующие категории ископаемых: субфоссилии, эуфоссилии, ихнофоссилии, копрофоссилии, хемофоссилии.
Субфоссилии (лат. sub — под, почти) представлены ископаемыми (почти ископаемыми), у которых сохранился не только скелет, но и слабоизмененные мягкие ткани. Для растительных остатков используют термин «фитолеймы» (греч. phyton — растение; leimma — остаток). Это в различной степени измененные растительные остатки, сохраняющие клеточную структуру. К субфоссилиям относятся фитолеймы из четвертичных отложений — семена, орехи, шишки хвойных, древесина, захороненные в торфяниках. Более измененные фитолеймы являются эуфоссилия-ми. К субфоссилиям принадлежат и уникальные находки некоторых животных этого времени, например мамонты, носороги и птицы. Консервантами для таких ископаемых являются вечная мерзлота, различные битумы, вулканические пеплы, эоловые пески. Считалось, что и янтарь является превосходным консервантом, однако в янтаре не сохраняются мягкие ткани. Вместе с тем ископаемые в янтаре (и растения и животные) полностью сохраняют свою форму, что позволяет тщательно изучить их внешнюю морфологию. Но попытка извлечь объект заканчивается тем, что все содержимое рассыпается в пыль. Значительно реже субфоссилии встречаются в отложениях более древних, чем четвертичные.
Эуфоссилии, или эвфоссилии (греч. еu — хорошо, настоящий) представлены целыми скелетами или фрагментами скелетов и их дискретными элементами, а также отпечатками и ядрами. Скелеты являются основными объектами палеонтологических исследований. Скелетные остатки имеют минеральный или органический состав. Это раковины и скелеты животных, оболочки бактерий и грибов, а также органические остатки листьев, семян, плодов, спор и пыльцы. Особо следует сказать о фитолеймах, представленных в различной степени обугленными остатками листьев, древесины, семян, плодов, спор и пыльцы.
Иногда используется термин «органикостенные микрофоссилии», к которым относятся оболочки бактерий и грибов, нитчатых цианобионтов, а также споры и пыльца. Размеры таких фоссилий менее 100 мкм. Органикостенные микрофоссилии могут иметь очень хорошую сохранность даже в докембрийских отложениях (см. рис. 250). Термин «органикостенные» можно распространять и на макроостатки.
Многие эуфоссилии сохраняют информацию не только о мягких частях организма и его функциональных системах, таких как кровеносная, половая, проводящие пучки растений и др., но и об образе жизни и биогеохимических процессах.
От скелетов и мягких частей организмов могут сохраняться отпечатки и ядра. Отпечатки представляют собой уплощенные оттиски, а ядра — объемные слепки полостей (рис. 1). Некоторые животные известны только по отпечаткам. Наиболее знаменитыми являются местонахождения отпечатков птиц, рыб, медузои-дов, червей, членистоногих и других животных, найденные в юрских золенгофенских сланцах Германии и в вендских отложениях Эдиакары Австралии. От растений чаще всего встречаются отпечатки листьев, реже стволов, семян и др. Отпечатки листьев отражают не только форму, но и характер жилкования. Отпечатки стволов сохраняют особенности поверхностного строения коры, например листовые подушки лепидодендроновых.
Среди ядер различают внутренние и внешние. Внутренние ядра возникают за счет заполнения породой внутренних полостей
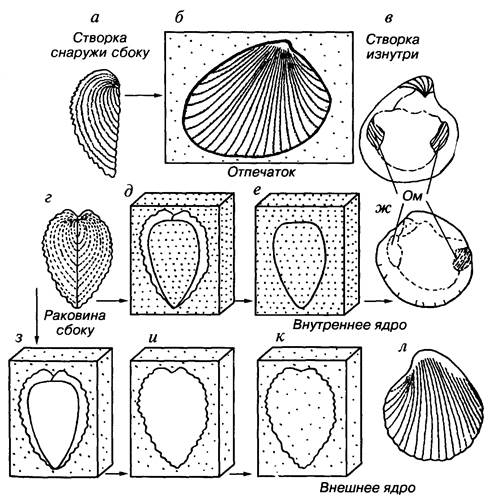
Рис. 1. Схема образования отпечатка (а-б), внутреннего ядра (в-ж) и внешнего ядра (з-л) на примере ребристой раковины двустворок. Ом — отпечатки мускулов
раковин двустворок, остракод, гастропод, брахиопод, аммонитов, а также черепных коробок позвоночных животных. Ядра растений чаще всего представляют отливы сердцевины стволов. Процесс возникновения внешних ядер сложнее, чем внутренних. Сначала скелет, заключенный в породе и ограничивающий полость, растворяется. Затем начинается заполнение породой вновь возникшей полости. Внутренние и внешние ядра наиболее четко отличаются друг от друга у скульптированных двустворок и брахиопод. На внутреннем ядре имеются отпечатки различных внутренних структур, а наружное ядро отражает особенности скульптуры раковины. Внешние ядра ребристые, шероховатые, грубые, а внутренние — гладкие, с отпечатками мускулов, связок и других элементов внутреннего строения.
Ихнофоссилии (греч. ichnos — след) представлены следами жизнедеятельности ископаемых организмов. Чаще всего они сохраняются в виде отпечатков, реже в виде слабообъемных образований. К ихнофоссилиям относят следы ползания и зарывания членистоногих, червей, двустворок; следы выедания, норки, ходы и следы сверления губок, двустворок, членистоногих; следы передвижения позвоночных.
Копрофоссилии (греч. kopros — помёт, навоз) состоят из продуктов жизнедеятельности ископаемых организмов. Они имеют объемный характер, сохраняясь в виде валиков, желваков, конкреций, холмиков, столбиков и даже пластовых тел. Термин «копрофосси-лии» был предложен в 1989 г. в учебнике «Общая палеонтология». За основу взято название «копролиты», введенное в научную литературу свыше 150 лет назад и обозначающее «окаменевшие экскременты животных» (Палеонтологический словарь, 1965).
К наиболее типичным копрофоссилиям относятся конечные продукты пищеварения илоедов и позвоночных животных; во второй группе копрофоссилий могут сохраняться непереваренные остатки других животных и растений. Копрофоссилии илоедов представлены валиками и ленточками, которые, на первый взгляд, как будто не отличаются от окружающей породы. Но, пройдя через кишечник илоеда, осадок обогащается кальцием, железом, магнием, калием и фосфором. В результате копрофоссилий илоедов приобретают более светлый или, наоборот, более темный, нередко красноватый оттенок, что и отличает их от окружающей породы. Процесс переработки осадка илоедами и биофильтраторами называют биотурбацией. Большинство осадочных отложений настоящего и прошлого проходят биотурбацию. Конечные продукты биотурба-ции и следы жизнедеятельности иногда называют биоглифами.
К копрофоссилиям можно отнести продукты жизнедеятельности бактерий и цианобионтов. Бактерии принимают участие в
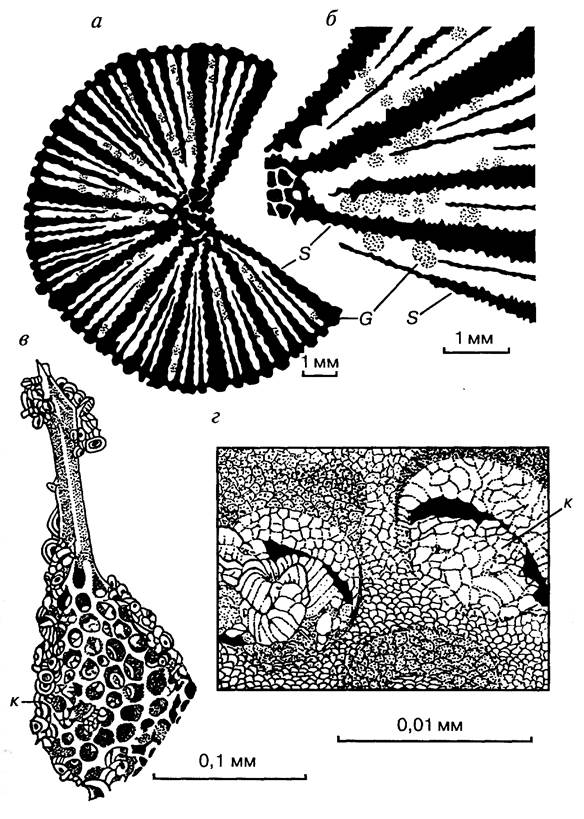
Рис. 2. Соотношения размеров скелетов различных групп организмов а, б — между септами коралла (макрофоссилии) застряли раковинки фораминифер (микрофоссилии); в-г — в «окошечках» скелета радиолярии (микрофоссилии) находятся кокколиты (нанофоссилии). G — Globigerinida, к — кокколиты, S — септы
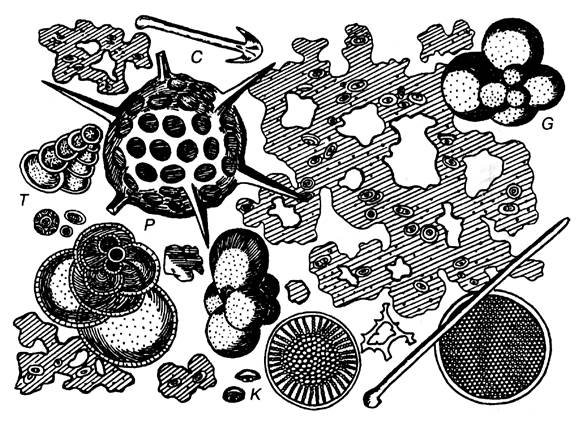
Рис. 3. Глубоководный органогенный ил (увел, в 700 раз), видна слизь с заключенными в ней кокколитами (К), раковинки Globigerinida (G) и Textulariida (T), a также радиолярии (Р) и спикулы губок (С) (Циттель, 1934)
образовании железистых, марганцевых и фосфоритовых конкреций, графитов, серы, нефти, газа и т. д. От жизнедеятельности цианобионтов сохраняются известковые слоистые образования — строматолиты, онколиты и катаграфии.
К хемофоссилиям (греч. chemie — химия) относят органические ископаемые биомолекулы бактериального, цианобионтного, растительного и животного происхождения. Хемофоссилии сохраняют химический состав биомолекул, позволяющий определить систематическое положение исходного организма, но не его морфологию. Изучение химического и таксономического разнообразия хемофоссилий тесно связано с проблемами возникновения и развития жизни, а также с происхождением горючих ископаемых, особенно нефти. Биологический фактор в формировании нефти долгое время отрицали, считая ее только хемогенной. Успехи в изучении хемофоссилий доказывают обратное. Хемофоссилии являются объектом изучения биохимии и молекулярной палеонтологии.
В зависимости от размеров ископаемых можно выделить (рис. 2 и 3): макрофоссилии (более 1 мм), микрофоссилии (десятые и сотые доли миллиметра) и нанофоссилии (сотые доли миллиметра и менее).
За фоссилии, кроме перечисленных форм сохранности, иногда принимают различные минералогические и литологические образования, получившие название псевдофоссилии (греч. pseudos — ложь). Разрозненные игольчатые минералы ошибочно описывают как спикулы губок, лучистые минеральные агрегаты — как колониальные кораллы, марганцевые дендриты — как растения, округлые желваки осадочного происхождения — как медузы и т. д.
ХИМИЧЕСКИЙ СОСТАВ И ПОРОДООБРА- ЗУЮЩАЯ РОЛЬ ИСКОПАЕМЫХ
Породообразующими называются ископаемые, которые составляют 30-40% и более от общего объема отложений. В образовании органогенной породы принимают участие как скелетные остатки, так и продукты жизнедеятельности, т. е. эуфоссилии и копрофоссилии. Иногда такие породы называют биолитами. Непременным условием породообразования является «скученный» характер обитания организмов. Таким свойством обладают в основном прикрепленные, малоподвижные и зарывающиеся формы, образующие заросли, банки, «луга», рифы и любые другие массовые поселения. Уже при жизни подобные организмы составляют основную часть биоценоза. Среди живущих в толще воды планктонных и нектонных организмов породообразующую роль играют те, которые обитают «стаями». Для континентальных организмов необходимы те же предпосылки. Органогенные породы могут возникать и за счет переноса первично единичных скелетов, образующих вторичные скопления в различных понижениях рельефа.
Состав скелетов ископаемых может быть минеральным, минерально-органическим и органическим. Среди минеральных скелетов наиболее распространены известковые, кремневые и фосфатные, образующие соответствующие по составу породы. Особенно много органогенных пород известкового (= карбонатного) состава (СаСО3): известняки, мергели, писчий мел, первичные и вторичные доломиты. Для названия органогенных пород, в отличие от хемогенных, используют прилагательное от латинского (греческого) названия той группы организмов, которая является основным породообразователем. Среди органогенных известняков и мраморов, образованных скелетами животных, наиболее распространены фораминиферовые, коралловые, строматопоровые, археоциатовые, губковые, серпулитовые, гастроподовые (птероподовые, спирателловые и др. ), цефалоподовые, остракодовые, мшанковые, брахиоподовые и криноидные. Известковые породы,
состоящие из скоплений раковинок двустворок, обычно называют ракушечниками (ракушняками), устричными горизонтами, рудистовыми известняками.
Скопления костей и скелетов позвоночных называют нейтрально — «костеносные слои». Растения, имеющие карбонатные скелеты, дают начало известнякам — водорослевым, литотамние-вым, харовым и кокколитовым (писчий мел). Органогенные известняки могут возникать и как конечные продукты жизнедеятельности цианобионтов и бактерий. От них остаются слоистые пластовые, желваковые, столбчатые, концентрические и «фигурные» образования — строматолиты, онколиты и катаграфии.
Для скоплений измельченных известковых скелетов и раковин различных систематических групп используют название «органогенный детрит». Обугленные остатки растений неопределимого систематического состава называют растительным детритом.
Минеральные скелеты кремневого (опалового — SiO2nH2O) состава встречаются реже, чем карбонатного. Они известны у одноклеточных животных (радиолярии, солнечники), у многоклеточных примитивных животных (губки), а также у низших одноклеточных водорослей (диатомовые и кремневые жгутиковые). Они образуют биогенные силициты. Кремневые породы — радиоляриты — состоят из скелетиков радиолярий, спонголиты — из спикул губок, диатомиты — из створок диатомовых водорослей. В каждой из перечисленных пород могут принимать участие кремневые скелеты других групп организмов. Так, в диатомитах иногда встречается небольшое количество радиолярий и спикул губок. А все они могут встречаться в таких биогенно-хемогенных породах, как трепел и опока.
Фосфатные скелеты в чистом виде встречаются редко, но фосфаты кальция (СаРО4 — в простейшем случае) как примесь или основная составляющая известны у многих организмов. Фосфаты кальция обнаружены у бактерий, грибов, книдарий (статолиты медуз и оболочки конулярий), червей, членистоногих, моллюсков, бра-хиопод, мшанок, иглокожих и позвоночных (конодонты, зубы акул, зубы и кости млекопитающих). Благодаря концентрации биогенного фосфата возникают месторождения фосфоритов. В Эстонии, например, источником биогенного фосфорита являются фосфорито-хитиновые раковины брахиопод ордовикского рода Obоlus s.l. (оболовые песчаники). В Подмосковье (например, в Лопатинском карьере) центрами фосфоритизации являются раковины позднеюрских аммоноидей. Фосфатная составляющая сконцентрирована в виде конкреций, желваков, оолитов и пластовых тел.
Минеральные скелеты всегда имеют органический матрикс, а также содержат элементы-примеси К, Mg, Ba, Si, Fe, Mn, Na, Ti,
F, Zn, Pb, Cu, Sr и др. В настоящее время обнаружены около 40 минералов, входящих в состав различных скелетов (табл. 1).
Кроме известковых и кремневых скелетов известны и такие экзотические, как целестиновые (SrSO4), встречающиеся у акантарий (одноклеточные). Несмотря на то что при жизни акантарии образуют скопления, они не стали породообразующими, так как их игольчатые скелетики после отмирания растворяются в воде, не попадая в осадок.
За счет жизнедеятельности бактерий образуются железистые, марганцевые, медистые и сульфидные месторождения, такие как железистые кварциты (джеспилиты) Кривого Рога, медистые песчаники Джезказгана. Бактерии участвуют в накоплении бокситов и фосфоритов.
Органические скелеты построены в основном углеводами, белками и жирами. Среди органических скелетов наиболее распространены хитиновые, хитиноподобные, шелкоподобные (спонгин), роговые и целлюлозные. Они образуют оболочки бактерий и циа-нобионтов, панцири, раковины и скелеты животных, а также ткани, оболочки спор и пыльцы растений. В органическом породообразо-вании самую большую роль играют высшие растения. Их массовые скопления при определенных процессах захоронения приводят к возникновению горючих ископаемых (каустобиолитов), таких как торф, уголь, горючие сланцы, нефть и газ. Происхождение нефти и газа связано с глубоким разрушением первичного органического состава, вызванным как жизнедеятельностью бактерий и цианоби-онтов, так и геологическими процессами. За счет жизнедеятельности высших растений образуются смолы (янтарь).
Необходимо отметить и рельефообразующую роль ископаемых и современных организмов, создающих рифовые постройки различного типа: береговые и барьерные рифы, атоллы, биостромы, биогермы и др. Рифовые постройки имеют сложное строение. Они состоят из комплекса взаимозамещающих известковых пород: органогенных, обломочно-органогенных и хемогенных.
Рифовые постройки возвышаются в рельефе в виде гряд, холмов и других поднятий. Их превышение над синхронными образованиями может быть довольно значительным. В образовании ископаемых и современных рифов принимают участие различные организмы (рис. 4). При восстановлении условий образования коралловых построек необходимо иметь в виду, что они могут быть и глубоководными. Иначе появляются ложные палеоэкологические и тектонические реконструкции. Основное отличие глубоководных от мелководных коралловых рифовых тел заключается в отсутствии растений и волновой эрозии. В результате цементом в глубоководных коралловых постройках служит известковый
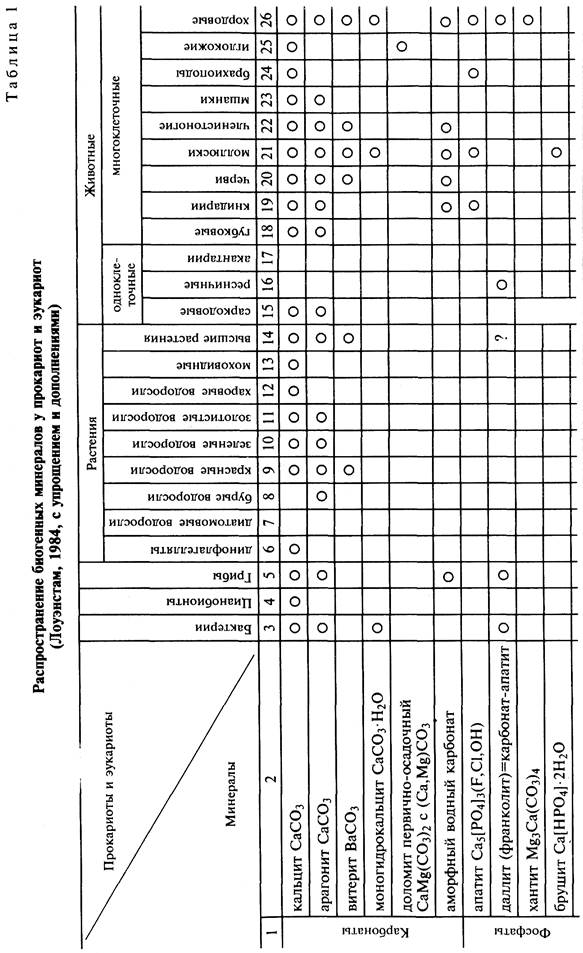
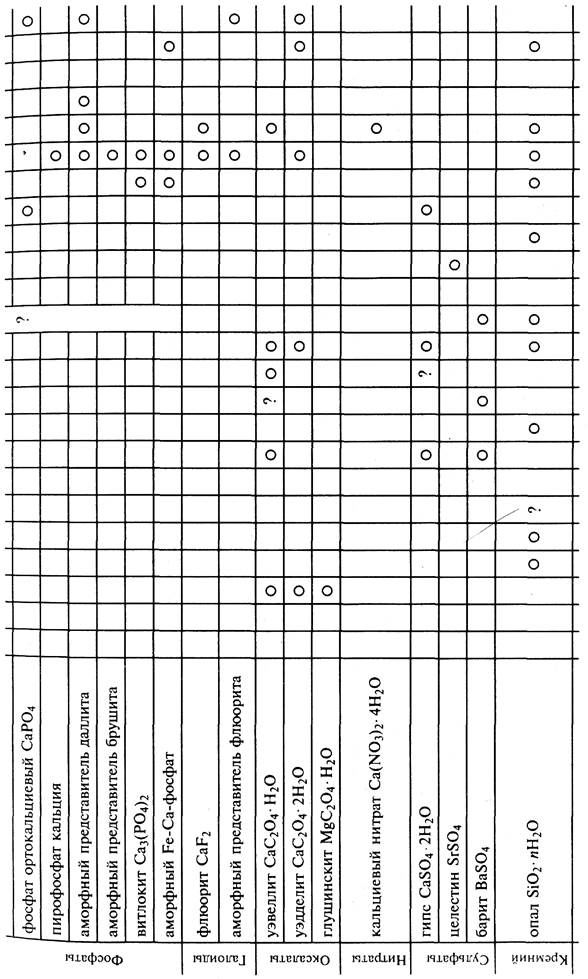
Рис. 4. Рифостроящие организмы прошлого и настоящего времени (Шеппард, 1987)
детрит животного происхождения, а в мелководных — детрит известковых водорослей и сами водоросли.
Ископаемые рифы служили коллекторами для нефти и газа, что позволяет рассматривать их в группе стратиформных месторождений. Кроме того, в ископаемых рифах могут накапливаться подземные пресные и минерализованные воды.
ИСТОРИЯ ПАЛЕОНТОЛОГИИ
Ископаемые, или окаменелости, человек использовал начиная с палеолита. Об этом свидетельствуют находки ожерелий из фрагментов вымерших кораллов, а также окремнелые панцири ископаемых морских ежей, использовавшихся в ритуалах погребений, и другие археологические находки. Различные ископаемые упоминаются в преданиях, мифах и сказках. Так, белемниты называют «чертовы пальцы», в восточных сказках их рассматривают как ногти джинов; раковины фораминифер-нуммулитид в сказаниях о битвах Александра Македонского описывают как окаменевшие монетки.
Первые научные письменные документы об ископаемых организмах принадлежат древнегреческим естествоиспытателям и
философам, и прежде всего Ксенофану (VI-V вв. до н. э. ) и Аристотелю (384-322 гг. до н. э. ). Ксенофан впервые описал ископаемые листья, у которых он находил сходство с современным лавром. Наиболее выдающийся философ-натуралист древности Аристотель разработал одну из первых классификаций органического мира. Окаменелости ему были известны благодаря находкам в осадочных породах Греции. Он считал их остатками морских животных, сохранившихся в тех местах, где раньше было море. Спустя много столетий в XV-XVI вв. такой взгляд на окаменелости поддерживал Леонардо да Винчи (1452-1519). Одновременно в среде ученых существовали иные воззрения на окаменелости как на объекты, созданные богом, оставшиеся после библейского потопа, или как на «игру» природы (воздействие лунного света и т. д. ).
В XVII—XVIII вв. начинаются интенсивные исследования в различных отраслях естествознания. Это привело не только к накоплению огромного фактического материала, но и к появлению различных идей, гипотез, методов и методик. Среди натуралистов того времени особое место занимает голландец А. ван Левенгук (1632-1723), которому принадлежит создание первого микроскопа. С появлением микроскопа перед учеными предстал новый мир неведомых ранее объектов и явлений. В XVIII в. практически одновременно работали трое блестящих ученых: в Швеции Карл Линней (1707-1723), во Франции Жорж Луи Бюффон (1707-1788) и в России М. В. Ломоносов (1711-1765).
Деятельность К. Линнея на протяжении нескольких десятилетий проходила в стенах Упсальского университета, в котором с 1741 г. он возглавлял кафедру ботаники. Помимо изучения растений, немало времени он посвятил и другим сторонам естествознания. Ему, в частности, принадлежат специальные исследования человекообразных. К. Линней ввел в научный обиход примерно 1000 терминов, большинство из которых было предложено им лично.
Этот выдающийся ученый знаменит прежде всего тем, что он лредложил правила и процедурные приемы для классификации разнообразия организмов и на их основании создал единую систему животных и растений. Свои первые представления об этом он изложил в работе «Система природы», которая вышла в 1735 г. и насчитывала всего 13 страниц. А спустя двадцать с лишним лет, в 1758-1759 гг., увидела свет новая работа, имеющая то же название, но состоящая из двух томов, каждый из которых насчитывал более 800 страниц. Вершиной научной деятельности К. Линнея стал труд «Philosophia Botanica», вышедший в Стокгольме в 1751 г. и изданный на русском языке в 1989 г. В этой работе К. Линней уже высказал твердое убеждение, что классификация растений
должна основываться на типе плодоношения, а в современном понимании — на строении органов размножения — фруктификаций (см. рис. 56, 57). Нельзя забывать, что во времена Линнея систематика растений строилась по самым различным признакам, включая даже алфавитный. Работы К. Линнея заложили основу Зоологического и Ботанического кодексов номенклатуры, имеющих статус международных законов для биологов и палеонтологов.
Разносторонние интересы и выдающуюся роль К. Линнея в развитии естествознания высоко оценили его современники как в Швеции, так и за ее пределами. В 1740 г. он стал первым президентом Академии наук в Стокгольме, в 1754 г. его избрали почетным членом Петербургской Академии наук, а в 1763 г. — членом Королевского общества в Лондоне. К концу жизни он получил дворянство и приобрел право писать свою фамилию de Linne.
С именем Линнея связано выделение многих родов беспозвоночных. Например, им установлены Tubipora, Isis (кораллы); Serpula (кольчатые черви); Patella, Buccinum, Helix (гастроподы); Arсa, Ostrea, Mytilus, Cardium, Pholas, Teredo (двустворчатые моллюски); Nautilus (головоногие) и т. д.
Научные исследования Ж. Бюффона нашли наиболее полное выражение в 44-томной «Естественной истории», издававшейся на протяжении 40 лет. Она принесла Ж. Бюффону мировую известность, что в немалой степени определялось блестящим и увлекательным стилем изложения.
Один из томов, носящий название «Эпохи природы», отражал взгляды Ж. Бюффона на историю Земли. Он считал, что окаменелости представляют остатки некогда живших животных и растений, а их разнообразие пытался объяснить их неодновременным существованием. Возраст Земли Ж. Бюффон оценивал в пределах от 6000 до 75 000 лет.
Начиная с 1739 г. Ж. Бюффон являлся директором Ботанического сада, основанного в 1626 г. королем Людовиком XIII и кардиналом Ришелье, а с 1640 г. получившего статус Королевского ботанического сада. При Людовике XV Ж. Бюффон получал достаточные средства на поддержание и расширение сада. Впоследствии многие французские натуралисты работали в этом ботаническом саду. Там же в ботаническом саду стоит памятник Ж. Бюффону с замечательной надписью: «Гению, по величию равному природе».
В России палеонтологические объекты стали специально собирать с начала XVIII в. для создания первого музея, известного под названием «Кунсткамера Петра Великого», послужившего прообразом большинства естественно-исторических академических музеев. В конце XVIII в. вице-президент Берг-коллегии А. Нартов
*** 22 ***
перевел с немецкого книгу «Валха Каменное царство». Вероятно, это было первое издание по палеонтологии на русском языке.
Основоположником естествознания в России стал М. В. Ломоносов, деятельность которого проходила в XVIII в. в царствование Анны Иоанновны, Елизаветы Петровны, Петра III и Екатерины II. Так же, как Аристотель и Леонардо да Винчи, М. В. Ломоносов считал, что ископаемые были обитателями прежних морских бассейнов. Причину гибели организмов М. В. Ломоносов видел в многократном чередовании водных и наземных условий, в результате чего захоронения морских организмов оказывались на суше. Неоднократное наступление и отступание моря он объяснял колебаниями «земной тверди». Во времена М. В. Ломоносова были известны находки «червяков и других гадин» в янтаре, и в работе «О слоях земных» он красочно описал, как они попали в смолу и окаменели. Возраст Земли М. В. Ломоносов оценивал в 400 000 лет. Надо сказать, что в XVIII в. подавляющее большинство ученых рассматривали ископаемых как остатки вымерших организмов. В это время актуальным стал и вопрос о развитии организмов, причинах их гибели и условиях захоронения.
Становление и развитие палеонтологии происходило в три этапа: додарвиновский, дарвиновский и последарвиновский. Этапы палеонтологии характеризуются господством того или иного мировоззрения. Так, в додарвиновский этап преобладало учение о неизменяемости видов, наиболее ярким выразителем которого был Ж. Кювье. Этому этапу предшествовало время первых описаний ископаемых организмов, выполненных в духе достижений и требований того времени. Например, у К. Линнея и в диссертации X. Фогта (1745), посвященной ископаемым кораллам Балтики, где впервые были изображены силурийские гелиолитиды. Однако уже в додарвиновский этап Ж. Сент-Илэр и Ж. Ламарк высказывали идеи об изменяемости видов.
Додарвиновский этап палеонтологии (конец XVIII — середина XIX в. ) связан с именами многих ученых, четверых из них считают родоначальниками основных научных направлений. Это английский инженер-геолог В. Смит (1769-1839; палеонтологический метод) и французские естествоиспытатели Ж. Б. Ламарк (1744-1829; палеозоология беспозвоночных), Ж. Кювье (1769-1832; палеозоология позвоночных), А. Броньяр (1801-1876; палеоботаника). Следует подчеркнуть, что изучение вымерших организмов с самого начала имело два аспекта: биологический (природа объекта) и геологический (нахождение ископаемых в определенной последовательности в земных слоях). Именно последняя особенность ископаемых в начале XVIII в. послужила основой для установления времени библейского потопа, а отсюда и возраста Земли.
*** 23 ***
Как индикаторы относительного возраста отложений ископаемые в самом общем виде были использованы профессором Горной академии в Саксонии А. Вернером (1750-1817), а позднее — детально, с внедрением в практическую деятельность — В. Смитом.
Инженер-геолог Вильям Смит в 1799 г. предложил два постулата: 1) слои, содержащие одинаковые окаменелости, являются одно-возрастными; 2) слои, содержащие разные окаменелости, являются разновозрастными (рис. 5). Эти два положения были столь просты и очевидны, что сразу вызвали понимание современников. На основании этих положений В. Смит построил сводный стратиграфический разрез Англии, где выделил 23 последовательных слоя, а также составил геологическую карту, где разновозрастные слои раскрасил различными цветами. Эти принципы Смита для построения сводного разреза и геологической карты уже на протяжении 200 лет являются классическими и незыблемыми.
Расчленение и корреляция отложений в биостратиграфии, основанные на двух положениях Смита, также остаются классическими, но с очень существенной оговоркой. Второе положение Смита, как оказалось впоследствии, не столь просто и однозначно, как первое. Разные окаменелости могут быть одновозрастными, но принадлежать разным экологическим обстановкам. Данное принципиальное уточнение стало учитываться последующими учеными. Эта поправка ничуть не умаляет достижений В. Смита, и его справедливо считают основателем палеонтологического
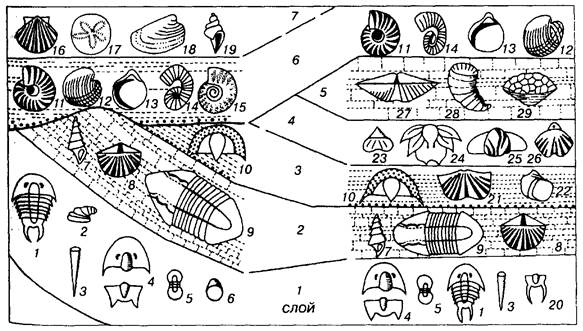
РИС. 5. Схема сопоставления двух разрезов по комплексам ископаемых (Moore, Lalicker, Fisher, 1952). Одинаковые ископаемые обозначены одинаковыми цифрами (1-29)
*** 24 ***
метода в геологии («метод Смита»). Таким образом, практическое применение ископаемых в геологии определило становление палеонтологии как науки.
Основы палеонтологии как науки были заложены почти одновременно тремя французскими натуралистами Ж. Б. Ламарком, Ж. Кювье и А. Броньяром.
Жан Батист Ламарк в молодости готовился к военной карьере, но в дальнейшем из-за состояния здоровья изменил свои планы и начал заниматься изучением растений. Его жизнь и работа на протяжении многих лет связаны с Королевским ботаническим садом Парижа, в котором в 1793 г. он получил место профессора естественной истории. После опубликования трехтомной сводки по флоре Франции (1778) Ламарк сконцентрировал свое внимание на зоологических объектах. Самыми знаменитыми трудами Ламарка являются «Система беспозвоночных животных» (1801), «Философия зоологии» (1809) и «Естественная история беспозвоночных животных» (1815-1822, 7 томов). Ж. Б. Ламарк не только развил идеи своих предшественников, в том числе Ж. Бюффона, о постепенных переходах между организмами, но и предложил новую трактовку истории органического мира.
Основные положения учения Ламарка: 1) виды существуют в течение определенного интервала времени и постепенно переходят один в другой; 2) общий путь истории развития органического мира — постепенное градуированное усложнение (развитие от низших к высшим) — принцип градации; 3) изменчивость видов, или приобретение новых признаков, связана с влиянием внешней среды и с упражнением или неупражнением органов (I закон Ламарка — закон изменчивости); 4) приобретенные изменения передаются потомкам и закрепляются через наследование (II закон Ламарка — закон наследования).
В истории естествознания Ламарк оставил очень глубокий след. Его главное достижение — создание первой эволюционной теории развития органического мира. Учение Ламарка получило название «ламаркизм» (по аналогии с дарвинизмом). Идеи Ламарка не получили поддержки у современников, многие считали его безумным. Ч. Дарвин в одном из писем писал: «Я почти убежден (в противоположность мнению, с которым я начал работу), что виды (это равносильно признанию в убийстве) не изменны... Выводы не так уж далеки от его (Ламарка) выводов, хотя способы изменений совершенно другие» (Избранные письма, 1950, с. 30). В настоящее время Ламарка считают одним из основателей учения об изменении видов и идейным предшественником Ч. Дарвина. Вопросы соотношения внешней среды, образа жизни, изменчивости и наследственности, поднятые Ламарком, до сих пор
*** 25 ***
актуальны. Ламарк начал подразделять животных на беспозвоночных и позвоночных, он также ввел термин «биология». Ламарк интенсивно изучал ископаемых беспозвоночных и включил их в «Учение о развитии органического мира».
С его именем связаны первые описания таких широко известных родов ископаемых беспозвоночных, как фораминиферы — Nodosaria, Lenticulina, Rotalia, Nummulites; кораллы — Cateniрога, Favosites, Alveolites, Sarcinula, Calceola, Cyclolites, Stylina, Fungia; гастроподы — Turritella, Ampullina, Lymnaea; двустворки — Nucula, Cucullaea, Plicatula, Gryphaea, Modiolus, Cyprina, Diceras, Hippurites, Radiolites; аммониты — Turrilites, Baculites; морские ежи — Clypeaster и т. д. Таким образом, вклад Ламарка в изучение современных и ископаемых беспозвоночных животных столь велик, что его по праву считают основателем этих разделов зоологии и палеонтологии.
Жорж Кювье начал систематически заниматься естествознанием с 1795 г., почти одновременно с Ламарком, а в 1802 г. стал профессором сравнительной анатомии в том же Ботаническом саду Парижа, где работал Ламарк. Научная деятельность Ж. Кювье вызывала у современников неизменное признание и восхищение. Историю органического мира Кювье рассматривал совершенно иначе, чем Ламарк. Он, как и К. Линней, считал, что виды неизменны и постоянны. Вместе с тем Кювье видел, что ископаемые разных слоев отличаются друг от друга. Он предложил такое объяснение: отличия ископаемых от слоя к слою связаны с периодическими революциями, после которых старое исчезает и возникает новое; постепенных переходов не существует. Это объяснение сразу приобрело многочисленных сторонников.
Кювье в своих рассуждениях об изменениях органического мира использовал слово «революция» (лат. revolutio — переворот, перестройка), реже его эквивалент «катастрофа» (греч. katastrophe — переворот, поворот). Впоследствии его объяснение развития органического мира стали называть теорией катастроф. В дальнейшем понятие «катастрофа» перестали связывать с неизменностью видов, а стали использовать его в различных сочетаниях, обычно как «неокатастрофизм» и «экологические кризисы» — для объяснения массовых вымираний, особенно значительных на рубеже ордовика и силура, палеозоя и мезозоя, мезозоя и кайнозоя.
Кювье был блестящим ученым, и его положительное научное наследие велико, особенно в области сравнительной анатомии. Он добыл и проработал огромное количество фактов, которые он согласно своему девизу «назвал, классифицировал и описал». Более того, на их основании Кювье вывел целый ряд закономерностей и законов. Кювье считал, что все особенности животного
*** 26 ***
подразделяются на главные и второстепенные. Первые следует использовать для выделения наиболее крупных единиц, вторые — для более мелких, подчиненных им таксономических единиц. Основой классификации и систематизации должно быть морфологическое сходство.
С именем Кювье связан закон корреляции органов, заключающийся в том, что органы и функции организма взаимосвязаны и составляют целостную систему (I принцип Кювье), соответственно приспособлен-ную к определенным условиям существования (II принцип Кювье). На основании закона корреляции он восстановил строение многих ископаемых позвоночных. Кювье ввел в биологию понятие «план строения», которому подчиняется разнообразие организмов. Он выделил четыре плана строения (позвоночные, моллюски, членистые и зоофиты, или радисты), предложив для них таксономическую единицу «тип», а также разработал общую систематику животных.
Кювье является основоположником палеозоологии позвоночных. Им описаны и реконструированы многочисленные ископаемые позвоночные, в том числе впервые установлены роды: Semnopithecus (обезьяны), Lagomys (зайцеобразные), Allactaga (тушканчиковые), Eligmodontia (хомяковые), Mephitis (скунсы), Mastodon (хоботные) и т. д. Кювье сгруппировал ископаемых и современных позвоночных в четыре фауны: 1) пресмыкающиеся, 2) палеотерии, анаплотерии, 3) млекопитающие мастодонты, мамонты, мегатерии, 4) современные виды плюс человек. Сравнительно-анатомические и палеонтологические работы Кювье, а также установленные им закономерности были тщательно учтены Ч. Дарвином и сыграли положительную роль в развитии эволюционного учения.
Этьен Жоффруа Сент-Илэр (1772-1844) — французский зоолог, последовательный эволюционист, современник Ламарка; коллега, соавтор, а затем идейный противник Кювье. В 1793 г., когда было принято решение Конвента о создании при Национальном музее естественной истории 12 кафедр, Жоффруа Сент-Илэр в возрасте 21 года получил предложение возглавить кафедру зоологии позвоночных. Спустя два года, в 1795 г., он пригласил на эту кафедру Жоржа Кювье и в том же году они издали две совместные работы, посвященные классификации млекопитающих и теории органов. Плодотворным явилось четырехлетнее пребывание в Египте, когда в 1798-1801 гг. Жоффруа Сент-Илэр в составе группы ученых сопровождал Наполеона. Позднее, в 1808 г., он был направлен в Португалию с заданием пополнить коллекции Национального музея Франции, а на следующий год возглавил кафедру зоологии в университете. Многие годы Жоффруа Сент-Илэр
*** 27 ***
трудился совместно с Кювье, завершение этих работ (1819-1837) нашло отражение в «Естественной истории млекопитающих».
Научное наследие Жоффруа Сент-Илэра многогранно. Ему принадлежат исследования в области сравнительной анатомии и эмбриологии; он создал теорию аналогов и разработал в противоположность Кювье учение о едином базисном плане организации всех типов животного мира; уделял большое внимание изучению уродств и аномалий, особенно их проявление на эмбриональных стадиях, считая, что это может привести к новым формам в более позднем возрасте.
В то же время основной причиной изменений в строении живых существ Жоффруа Сент-Илэр, подобно Бюффону и Ламарку, считал изменение внешней среды. В историю естествознания он вошел прежде всего как последовательный эволюционист, выступивший с критикой взглядов Кювье. Уже в 1802 г. Жоффруа Сент-Илэр и Ламарк заложили основные положения эволюционной теории.
В 1830 г. в Парижской Академии наук состоялся публичный диспут, на котором Кювье критиковал Жоффруа Сент-Илэра за единый план строения и причинное толкование явлений, в то время как его противник выступал против четырех изолированных типов строения животного мира Кювье, а главное, отстаивал эволюционный взгляд на развитие живой природы. Этот научный спор между трансформистами и катастрофистами завершился победой Кювье, и на долгие годы во Франции наступил период господства идеи о неизменяемости видов и объяснения изменения живой природы катастрофическими преобразованиями. Спустя 30 лет, когда был опубликован труд Ч. Дарвина (1859), именно во Франции, где ранее работали такие блестящие эволюционисты, как Бюффон, Ламарк и Жоффруа Сент-Илэр, идеи Дарвина с большим трудом находили своих сторонников.
После нашумевшего диспута 1830 г. Кювье прожил всего два года. Многолетний совместный труд о млекопитающих, о чем говорилось выше, Жоффруа Сент-Илэр завершил без него.
Адольф Броньяр на протяжении всей жизни целенаправленно занимался изучением растений. Его основные труды посвящены кайнозойской растительности. С его именем связано становление палеоботаники, т. е. палеонтологии растений. Броньяр предложил первую единую систематику растений, проследил и описал историю развития растений прошлых геологических эпох (1828-1838).
Ламарк, Кювье и Броньяр использовали ископаемых для определения относительного возраста вмещающих отложений и реконструкций условий обитания (глубина бассейна, палеоклимат). Они также давали оценку продолжительности существования Зем-ли как планеты.
*** 28 ***
Додарвинский период становления палеонтологии в России связан прежде всего с именами X. И. Пандера, П. М. Языкова и Г. И. Фишера фон Вальдгейма. X. Пандер палеонтологически обосновал стратиграфию кембрия и ордовика окрестностей Петербурга и сделал 200 рисунков ископаемых (1830). Он открыл также новую группу микроскопических ископаемых — конодонты, которые приобрели чрезвычайно важное стратиграфическое значение в современных исследованиях палеозоя. П. М. Языков (1832) на основании изучения конкретных разрезов и распределения в них ископаемых впервые разработал стратиграфию юры и мела Поволжья. Он ввел в научную литературу русские эквиваленты иностранным терминам и латинским названиям, такие как ярус, брюхоногие, головоногие моллюски, плеченогие.
Г. И. Фишер являлся профессором Московского университета и занимался биостратиграфией и фауной палеозоя и мезозоя Подмосковья (1837). Он впервые описал такие известные ископаемые роды, как фораминиферы — Fusulina, Streblus; кораллы — Chaetetes, Halysites; двустворки — Amphidonta; брахиоподы — Choristites и т. д. Фишер организовал при Московском университете Московское общество испытателей природы — МОИП (1806) и был его первым президентом. Учрежденное им в начале прошлого века общество и сейчас активно продолжает свою деятельность. При обществе была создана библиотека, функционирующая и в настоящее время. Она содержит уникальные старинные издания по естествознанию, в том числе и первую книгу по палеонтологии на русском языке. Книга была опубликована по поручению Екатерины II и посвящена ей.
Первые два учебника были изданы в России в начале XIX в.: по палеозоологии беспозвоночных — «Конхиология... » (греч. соnche — раковина) и палеоботанике — «Сокращенное руководство к систематическому определению ископаемых растений... », написанные Я. Г. Зембницким (1831, 1833). Зембницкий перевел многие латинские и латинизированные термины и названия родов и видов, приведя для них русские эквиваленты (например, Lepidodendron — чешуедрев, Sphenopteris — клинокрыл).
Постепенно изучение ископаемых в России в XIX в. приобрело биологический характер, но биостратиграфическая направленность при этом обязательно сохранялась (К. Ф. Рулье, Э. И. Эйхвальд и др. ). К. Ф. Рулье (1814—1858) был сторонником изменяемости видов. Изучая ископаемых Подмосковья, он подчеркивал неразрывность истории развития живой и неживой природы. Рулье дал впервые палеоэкологические и палеоклиматические реконструкции юры Подмосковья. Э. И. Эйхвальд (1795-1876) описал коллекцию ископаемых России, включающую беспозвоночных, позвоночных
*** 29 ***
и растения всего фанерозоя. Он издал книгу под названием «Палеонтология России» на русском и французском языках (1860-1868). Экспозиция коллекции Эйхвальда, явившаяся первым единым собранием ископаемых России, была развернута в Петербургском университете, где она находится и в настоящее время. Ценность коллекции была ясна уже современникам, недаром миллионер Дж. Пибоди хотел приобрести ее для американского музея.
Дарвиновский этап палеонтологии (середина — конец XIX в. ). Теоретические и практические достижения палеонтологии как науки тесно связаны с общими успехами биологии. К середине XIX в. биологи установили ряд закономерностей в строении и развитии живого, в том числе (кроме упомянутых выше): отсутствие резкой границы между животным и растительным миром (Ж. Бюффон); клеточное строение, становление зародышевых листков (X. Пандер); закон соответствия зародышей высших и низших животных и закон о последовательности закладки признаков в эмбриогенезе от общих к частным (К. М. Бэр); значение изменчивости и наследственности в выведении новых разновидностей растений и животных и т. д.
Чарльз Дарвин (1809-1882) родился в семье врача. В годы учебы в гимназии у него проявилась любовь к природе и коллекционированию минералов, растений, насекомых, раковин и пр. Два года занятий на медицинском факультете Эдинбургского университета (1825-1827), а затем на богословском факультете в Кембридже не завершились выбором профессии. Пересилил интерес к живой природе, совместные экскурсии со знакомыми натуралистами, наблюдения над животными и растениями. Большое влияние на дальнейшую судьбу Ч. Дарвина оказал профессор ботаники Кембриджского университета Д. Генсло, который был учителем и другом Ч. Дарвина. К планомерным работам по естествознанию Ч. Дарвин приступил в 1831 г., когда по рекомендации Генсло был принят в качестве коллектора на научно-исследовательский корабль «Бигль» для участия в кругосветном путешествии. Материалы пятилетнего плавания определили направление всех дальнейших работ Дарвина и легли в основу его учения, названного впоследствии теорией эволюции, или дарвинизмом.
Дарвин установил, что эволюция, т. е. процесс исторического развития органического мира, может происходить только при сочетании трех непременных условий: изменчивости, наследственности и естественного отбора. Эта триада условий является целостным движущим фактором эволюции. Так, если предположить, что отсутствует изменчивость, то естественный отбор будет проявлять себя альтернативно: или все уничтожать, как неприспособленное к существованию, или все сохранять, но до определенного
*** 30 ***
времени, пока не изменятся условия. В любом случае развития не происходит. В действительности наблюдается непрерывное развитие органического мира, выражающееся в появлении и вымирании различных групп организмов, образующих пространственные и временные биоценозы и биоты, определяющие развитие биосферы в целом.
Полный вариант теории Дарвина был опубликован в 1859 г. в его книге «Происхождение видов». Для разработки эволюционного учения он привлек и палеонтологический материал. Выводы, к которым пришел Дарвин, многочисленны и разнообразны; укажем следующие: 1) если вид однажды исчез, то он не возникнет вновь (закон о необратимости эволюции); 2) более специализированная организация потомков, лучше приспособленная к окружающей среде, приводит ко все более усложненным формам (причина прогрессивной эволюции); 3) определенные организмы могут существовать в определенных условиях, поэтому эволюция является по своей сути приспособительной, скоррелированной с условиями обитания (адаптивная эволюция); 4) расхождение, или дивергенция, признаков происходит путем расщепления признаков от одной предковой формы (монофилетическая эволюция); 5) в ископаемом состоянии сохраняется незначительное число организмов (неполнота геологической летописи).
Теория Дарвина явилась мощным импульсом к дальнейшим исследованиям в этом направлении, позволившим обосновать, детализировать и раскрыть ряд неизвестных ранее закономерностей, а некоторые заключения в дальнейшем пересмотреть. Но незыблемым осталось основное содержание эволюционной теории, заложенной Дарвиным: развитие органического мира осуществляется через триаду — изменчивость, наследственность, отбор. Непосредственно после выхода в свет «Происхождения видов» самым настойчивым пропагандистом и защитником теории Дарвина стал Т. Г. Хаксли (Huxley, 1825-1895). И это несмотря на то, что ранее Хаксли придерживался взглядов Линнея и Кювье о неизменяемости видов. К научным заслугам Хаксли следует отнести: установление родственных связей между медузами и полипами, доказательство происхождения птиц от пресмыкающихся, опровержение гипотезы, в соответствии с которой череп позвоночных произошел от видоизмененных позвонков, и ряд других научных положений.
Дарвинский этап развития палеонтологии характеризуется фактическим обоснованием теоретических положений эволюционной палеонтологии. Во второй половине XIX в. трудилась блестящая плеяда естествоиспытателей: В. Вааген, М. Неймар, В. О. Ковалевский, Л. Долло и др. Особое внимание было обращено на
*** 31 ***
послойное изучение ископаемых, что позволило построить филогенетические ряды развития вымерших животных. Одна из таких работ была проведена В. Ваагеном (Waagen, 1869) по юрским аммонитам; он различал изменчивость организмов в пространстве (разновидность) и их изменения во времени (мутации, термин Ваагена). Смена мутаций, как считал Вааген, может служить основой зональной шкалы.
Эволюционную теорию Ч. Дарвина поддержал и подкрепил собственными исследованиями венский палеонтолог и геолог М. Неймайр (1845-1890). Он показал изменяемость видов на раковинах брюхоногих моллюсков, последовательно собранных из неогеновых отложений Западной Славонии (Югославия). Временной ряд форм рода Paludina отчетливо иллюстрирует как унаследованность формы и скульптуры при переходе от одной формы к другой, так и резкое отличие крайних членов этого ряда (рис. 6). Причиной изменчивости М. Неймайр считал влияние окружающей среды. «Ни в одной области естествознания учению о происхождении видов не суждено оказать большего влияния, как в палеонтологии, которая призвана дать историю органической жизни и наблюдать постепенное развитие ее» (Неймайр, 1898, с. 6).
Одним из первых русских ученых, который воспринял учение Ч. Дарвина, был В. О. Ковалевский (1842-1883). Для подтверждения дарвиновской теории он использовал палеонтологические данные. Объектом исследования В. О. Ковалевского явились ископаемые копытные, при изучении которых затрагивались проблемы морфофункционального анализа скелетных остатков, выявлялись связи между строением скелета животного со средой обитания, закладывались основы науки, позднее названной «палеоэкология». В. О. Ковалевский разработал новую классификацию копытных, доказывая их происхождение от общего предка (принцип радиации). Во всех его исследованиях вопросы о факторах эволюции рассматривались с точки зрения естественного отбора. В. О. Ковалевского считают одним из основателей эволюционной палеонтологии.
Рис. 6. Последовательный ряд мутаций раковин гастропод (а-е) из палюдиновых слоев неогена Западной Славонии (Неймайр, 1903, с упрощением)
*** 32 ***
В области палеонтологии позвоночных последователем В. О. Ковалевского был бельгийский исследователь, убежденный дарвинист Л. Долло (1857-1931). Одна из его работ — «Законы эволюции» (1893) — содержала теоретические обобщения, развивавшие эволюционную палеонтологию и подкреплявшие учение Дарвина. Л. Долло сформулировал мысль о прерывистости эволюции, т. е. о наличии в ходе развития резких скачков. Свою мысль он пояснил следующим образом. Увеличение длины тела позвоночных может идти как за счет вытягивания отдельных позвонков, так и за счет возрастания числа позвонков. Во втором случае постепенность исключена, возможно лишь скачкообразное развитие. Вторая высказанная Л. Долло закономерность сводится к тому, что организм после прохождения определенного цикла непременно вымирает. Широкому кругу естествоиспытателей Л. Долло известен как автор закона о необратимости эволюции, т. е. о невозможности возврата потомков к состоянию, характерному для предков. Впоследствии появились факты, указывающие на возможность частичного возврата предковых структур у некоторых потомков.
После организации в 1882 г. Геологического комитета, во главе которого стал выдающийся геолог и палеонтолог А. П. Карпинский, было начато систематическое изучение геологического строения территории России. Оно сопровождалось тщательным изучением ископаемых. Исследования на юге России и в Сибири положили начало накоплению отечественного материала по ископаемым позвоночным.
Последарвиновский этап палеонтологии (XX в. ). Достижения в развитии эволюционного учения в XX в. многообразны и значительны. Следует только иметь в виду, что триада Дарвина — изменчивость, наследственность, естественный отбор — осталась базисной при разработке любого эволюционного учения.
Появление новых теорий эволюции опирается на исследования по генетике, молекулярной биологии, цитологии, биохимии, биофизике, эмбриологии, экологии, популяционной биологии и т. д. Новые теории эволюции объясняют конкретные механизмы, пути, формы и темпы проявления всей триады Дарвина или одного-двух ее факторов нередко на примере какой-нибудь определенной группы органического мира.
Новые объяснения не всегда совпадают с разъяснениями Дарвина. Так, русский ученый Л. Берг (1922) утверждал, что эволюция идет на основе закономерностей (номогенез), а не случайностей, как полагал Дарвин. Из многих положений теории номогенеза Берга отметим следующие: 1) эволюция идет через изменение почти всего состава особей, а не трансмутации его отдельных представителей; 2) онтогенез и филогенез протекают по одним и тем
*** 33 ***
же законам, а именно: одни признаки «повторяют прежние стадии, Другие предваряют будущие»; 3) развитие органического мира не является монофилетичным.
Морфологические закономерности эволюции были тщательно проанализированы, описаны и классифицированы русским биологом А. Н. Северцовым (1931, 1939, 1945 и др. ). Ему принадлежит дальнейшая разработка теории филэмбриогенеза. С именем А. Н. Северцова также связано раскрытие путей и способов биологического и морфофи-зиологического прогресса и регресса.
В создании синтетической теории эволюции принимали участие ученые многих стран, но ее крестным отцом считают американского палеонтолога Дж. Симпсона (1944, 1948, 1953). Синтетическая теория эволюции по существу базируется на универсальной триаде Дарвина, но конкретизированной и детализированной с позиций генетики и популяционной биологии. По Симпсону, основными факторами эволюции являются следующие семь: воздействие внешней среды, мутагенез, строение популяции, скорость смены поколений и их продолжительность, колебания численности, естественный отбор, репродуктивная изоляция. Целенаправленный (векторный) характер эволюции по синтетической теории обусловлен естественным отбором. Следует отметить, что перечисленные факторы тесно взаимосвязаны и взаимообусловлены, но в зависимости от реальных обстоятельств нередко кажется, что какие-то из них важнее других.
Во второй половине XX в. очень плодотворной оказалась идея использовать достижения кибернетики для объяснения многих явлений эволюции. В России кибернетические проблемы эволюции разработаны И. Шмальгаузеном и его школой (1968). Шмальгаузену принадлежит постановка вопроса об изменяющихся факторах эволюции (1966, 1974), а также разработка теории стабилизирующего и дестабилизирующего (преобразующего) отбора.
Развитие эволюционной палеонтологии тесно связано не только с успехами в биологии, но и с интенсивными геологическими исследованиями. Стала актуальной проблема подготовки специальных кадров. Во многих странах мира еще до второй мировой войны были созданы кафедры палеонтологии и соответствующие ИМ институты, стали выходить палеонтологические журналы и Другие периодические издания. Возникла необходимость систематизировать весь фактический материал по палеонтологии. Три страны: Франция, Советский Союз и Соединенные Штаты Америки издали соответствующие сводки: «Traite de Paleontologie» (З тома, 1952-1953), «Основы палеонтологии» (15 томов, 1958-1964), «Treatise on Invertebrate Paleontology» (более 20 томов, 1953-1981). После ревизии отдельные тома американской сводки переиздаются.
*** 34 ***
Одним из первых крупных организаторов палеонтологической науки и университетского палеонтологического образования в послереволюционной России был А. А. Борисяк (1872-1944). Научная деятельность А. А. Борисяка в области палеонтологии была посвящена изучению различных групп палеогеновых — четвертичных млекопитающих: копытных (носороги, «индрикотериевая» фауна Казахстана, лошади, халикотерии), хоботных, пещерных медведей, моржей и других животных из разных районов страны. На протяжении всей жизни А. А. Борисяк последовательно отстаивал взгляд на палеонтологию как на самостоятельную биологическую науку, а не только как на «служанку геологии» (палеонтологический метод), что нашло отражение в его научной, научно-организационной и учебно-организационной работе. В различные периоды жизни он трудился в Геологическом комитете, преподавал на кафедре палеонтологии (1898-1899), а позднее на кафедре исторической геологии (1911-1930) Ленинградского горного института, основал Палеонтологический институт АН СССР и руководил им (1930-1944) и кафедрой палеонтологии Московского университета (1939-1944). Педагогическая работа А. А. Борисяка неразрывно связана с созданием учебников по палеонтологии и исторической геологии, получивших широкое признание и выдержавших несколько изданий.
Ю. А. Орлов (1893-1966), достойный преемник А. А. Борисяка, был выдающимся ученым и крупнейшим организатором науки в Советском Союзе. С его деятельностью связаны дальнейшее развитие Палеонтологического института Академии наук и кафедры палеонтологии Московского университета, организация нового Палеонтологического музея, носящего ныне его имя, издание 15 томов «Основ палеонтологии» (1958-1964), учебника «Палеонтология беспозвоночных » (1962), основание «Палеонтологического журнала» (1958), организация многих крупных экспедиций, в одной из которых он открыл третичные фауны на реках Ишим и Иртыш. Научные интересы Ю. А. Орлова были связаны с палеоневрологией. Он показал, что у ископаемых животных по характеру развития разных долей мозга можно оценить степень обоняния, слуха (височные доли), интерпретацию звука (лобные доли), зрение (затылочная доля), ловкость и быстроту движений (полушария мозжечка). В своих работах Ю. А. Орлов также доказал, что слепки черепной коробки не полностью совпадают с объемом, формой и рельефом мозга, нередко они отражают также строение и внутреннюю скульптуру черепной коробки.
*** 35 ***
РАЗДЕЛЫ ПАЛЕОНТОЛОГИИ
Разделы палеонтологии отражают различные аспекты изучения ископаемых, а конечной целью всех разделов является реконструкция развития органического мира. Это определяет главную задачу палеонтологии как науки. Знание законов развития жизни приобретает все большее значение в связи с пониманием роли биосферы в развитии планеты. Прикладное использование палеонтологии связано с геологией, где палеонтологический метод определения относительного возраста пород до сих пор является основным для фанерозоя. Разделы палеонтологии тесно связаны друг с другом. Успехи (и ошибки) одного направления прямо или косвенно влияют на исследования и результаты других направлений.
Палеонтология (палеозоология) беспозвоночных занимается изучением всех ископаемых животных, за исключением хордовых. Беспозвоночные были выделены Ж. Б. Ламарком в начале XIX в. как систематическая категория Invertebrata в противоположность позвоночным — Vertebrata. В дальнейшем оказалось, что позвоночные являются частью типа хордовых и поэтому нельзя противопоставлять их как одноранговую категорию многочисленным типам беспозвоночных. Тем не менее термин «беспозвоночные» сохранился до сих пор как удобный и понятный. Среди ископаемых беспозвоночных известны следующие типы: саркодовые, ресничные, губковые, археоциаты, книдарии, гребневики, черви (кольчатые, плоские, приапулиды и др. ), моллюски, членистоногие, мшанки, брахиоподы, иглокожие, полухордовые, погонофоры, конодонтофориды.
Палеонтология беспозвоночных изучает биологическое разнообразие и его изменение во времени и в пространстве. Биологическое разнообразие устанавливают через морфологию, определение систематического состава и построения новых классификаций и систем. Эта работа сопровождается изучением изменчивости и морфогенеза, морфофункциональным анализом, а также другими исследованиями, связанными с содержанием разнообразных разделов палеонтологии.
Палеонтология (палеозоология) позвоночных изучает тип хор-довых, преимущественно один из его подтипов — позвоночные. Основоположником раздела является Ж. Кювье (начало XIX в. ). Последовательность изучения и круг вопросов те же, что и в палеонтологии беспозвоночных.
Палеоботаника исследует ископаемые растения. Основоположником раздела был А. Броньяр (начало XIX в. ). Определение систематического разнообразия растений более затруднено, чем животных, оттого, что разные части растений сохраняются обычно
*** 36 ***
раздельно. Они получают самостоятельные названия как представители разных организмов, тем самым искажая исходное систематическое разнообразие. Исследование спор и пыльцы (палинология, или спорово-пыльцевой анализ) показывает большее систематическое разнообразие, чем изучение других остатков растений. Грибы, минеральные продукты жизнедеятельности бактерий и цианобионтов, а также оболочки бактерий до последнего времени рассматривали в разделе «Палеоботаника». Палеоботаника, как и палеозоология, кроме вопросов морфологии, систематики и реконструкции растений занимается историческими, тафономическими, палеогеографическими и другими исследованиями.
Раздел «Палеонтологические проблематики» или просто «Проблематики» рассматривает морфологию скелетов организмов неясного систематического положения с обязательным морфофункциональным анализом и указанием возможных родственников. Обособление «проблематик» в самостоятельный раздел связано с многочисленными дискретными микроскопическими остатками, выявленными преимущественно благодаря химическому препарированию (вторая половина XX в. ). Проблематики микроскопических размеров представлены акритархами, хитинозоа, склеритами кембрийских организмов и т. д. Из макрофоссилий к проблемати-кам относят: афросальпингидеи, рецептакулиты, вольбортеллиды, стенотекоидеи и др. Перечисленные проблематики иногда предлагают называть «загадочными животными», «загадочными организмами» или «загадочными ископаемыми», так как их биологическое происхождение очевидно, а термином «проблематики» принято обозначать находки, биологическая сущность которых сомнительна и не доказана. Систематическое и морфотопографическое положение дискретных остатков загадочных групп иногда удавалось восстановить после находок макрофоссилий сравнительно полной сохранности. Так произошло после открытия конодонтоносителя. Находки организмов в среднем кембрии Британской Колумбии показывают, что различные загадочные «чешуи» могут принадлежать одному организму, а не разным (рис. 7).

Рис. 7. Морфология «чешуи» животного неясного систематического положения. Канада, Британская Колумбия. Сланцы Бёрджес, средний кембрий (Conway Morris, Whittington, 1985)
*** 37 ***
Микропалеонтология изучает микро- и нанофоссилии. К ним принадлежат не только мелкие и мельчайшие организмы, но и части крупных животных и их стадии развития, имеющие микроскопическую размерность. Таким образом, объектом микропалеонтологии могут быть любые остатки организмов, требующие для своего изучения технических средств в виде луп и микроскопов. Но объектами преимущественного изучения стали немногие: среди животных — фораминиферы, радиолярии, тинтинниды, остракоды, конодонты, чешуи телодонтов, зубы мелких млекопитающих, отолиты рыб; среди низших растений — диатомовые водоросли, кремневые жгутиковые, кокколитофориды; среди высших растений — споры и пыльца; среди проблематик — акритархи и хитинозоа. Обособление микропалеонтологии как особого раздела связано с началом интенсивного бурения на нефть и газ в 30-х годах XX столетия. Микро- и нанофоссилии в керне скважин многочисленны, они отвечают условию «в малом объеме — максимум информации», в то время как макрофоссилии в керне фрагментарны и редки.
Раздел палеонтологии «Докембрийская биота» приобрел самостоятельность во второй половине XX в. после открытия эдиакар-ской бесскелетной фауны в Австралии, отвечающей по времени венду (= терминальный рифей). Вендскую фауну с самого начала стали изучать как биоту, т. е. как совокупность организмов определенного временного — пространственного отрезка. Понятие «докембрийская биота» сейчас шире, оно также включает организмы археозоя и раннего-среднего протерозоя.
Палеоэкология изучает среду обитания ископаемых организмов, которая складывается из взаимоотношений друг с другом (биотическая среда) и с неживой природой (абиотическая среда). Палеоэкология исследует как отдельные органи
Дата добавления: 2015-06-27; просмотров: 6399;