Пассивный транспорт и его виды
Подчиняется закону Фика:
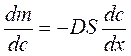
Скорость переноса вещества зависит от градиента концентрации – dc/dx, площади (S) и коэффициента (D) диффузии.
Подразделяется на:
1. Простую диффузия.
2. Облегченную диффузия.
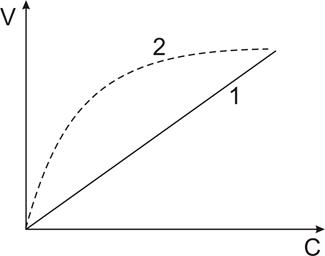
| Рис. 11.Зависимость скорости пассивного транспорта от концентрационного градиента: 1- Простая диффузия; 2.- облегченная диффузия. |
Простая диффузия отличается отсутствием специальных систем для переноса веществ. Подчиняется правилам Овертона: Скорость проникновения через мембрану прямо пропорциональна липотропности вещества и обратно пропорциональна – его размерам.
Позднее Колландер из уравнения Фика вывел более удобно выражение для расчета потока (J) поступления веществ в клетку:
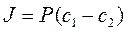 , где:
, где:
с1 и с2- разность концентраций по обе стороны мембраны
Облегченную диффузия отличает наличие специфических структур, способствующих транспорту через мембрану:
а) переносчики;
б) ионные каналы.
Наличие переносчиков изменяет кинетику транспорта, и она становится сходной с уравнениями ферментативного катализа, только в роли фермента выступает переносчик, в роли субстрата переносимое вещество (S):
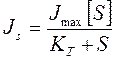 ,
,
где КT- константа транспорта соответствует константе Михаэлиса и равна концентрации S при Js=Jmax/2.
Представители систем облегченной диффузии ионов (симпорт, антипорт) с участием специфических переносчиков:
| Na+/Cl—обмен Na+/Na+--обмен Na+/H+ – обмен Na+/Ca2+-обмен Cl-/HCO3-+-обмен Cl-/ Cl--обмен | 
| антипорт – котранспорт |
K+/Cl—и Na+,K+,2Cl- – симпорт – котранспорт
Унипорт
Ионофоры-небольшие гидрофобные молекулы, которые растворяются в липидном слое мембран и увеличивают проницаемость мембран для ионов. Большинство ионофоров синтезируется бактериями (в качестве оружия против своих конкурентов), некоторые являются антибиотиками. Широко используются в клеточной биологии.
Ионофоры делятся на:
1. Подвижные переносчики. Имеют кольцеобразную структуру: наружняя часть – гидрофобная и контактируют с липидным слоем; внутренняя часть – гидрофильная содержит карбоксильные атомы кислорода.
2. Каналообразующие. Состоят из линейных полипептидов, которые имеют гидрофобные боковые цепи. Две таких молекулы образуют канал
Ионные каналы (унипорт) класссифицируют:
А) по типу ионов: натриевые, калиевые, кальциевые и хлорные –каналы;
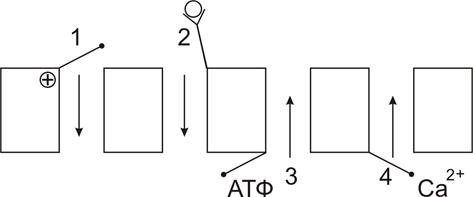
Б) способу регуляции (Рис.12):
1. потенциал-чувствительные
2. хемочувствительные (рецептор-управляемые)
3. внутриклеточными веществами (ионами, ФАВ).
|
| Рис.12. Классификация ионных каналов по способу регуляции. 1-потенциал-чувствительные; 2-хемочувствительные (рецептор-управляемые); 3-АТФ–чувствительные; 4-кальций-зависимые. |
Натриевые и калиевые каналы
Размеры ионных каналов меньше одного нМ (02-04 нМ). Эти структуры обеспечивают важнейшие электрофизиологические свойства клеток и тканей, обеспечивая прохождение в 1с 106-109 ионов, регулируются мембранным потенциалом и БАВ. Через ионные каналы Na+ и K+ следуют совместно с одной молекулой воды-гидратной оболочкой, Хотя в растворе таких оболочек может быть несколько (до 10). Таким образом, при прохождении через ионный канал происходит дегидратация катиона, на что требуется энергия. Роль этой энергии выполняет градиент ионов, так как ионный транспорт относится к пассивному виду переноса веществ через мембрану. В процессе переноса катионов должны выполнятся два основных условия (фактора):
1. Стерический – совпадение размеров катиона+гидратная оболочка с размерами канала.
2. Энергетический – взаимодействие катиона с карбоксильными (ОТРИЦАТЕЛЬНО заряженными группировками самого канала).
Кальциевые каналы
Кальциевые каналы делятся на потенциал-зависимые и рецептор–управляемые. Они находятся в плазменной мембране всех электровозбудимых клеток: сердечная и гладкая мышца, мышечные волокна членистоногих, нервная клетка (соматическая, дендрита), эндокринные и иммунные клетки и т.д.
В нормальных условиях внутри клетки [Ca2+]in = 10–7 – 10–6 M
Через кальциевый канал проникают только двух валентные катионы в соответствии с рядом:
pKBa2+ > pKSr2+ > pKCa2+ > pKCo2+ > pKNi2+ > pKCd2+
Из этого ряда следует, что эти катионы в отношении их взаимодействия с кальциевым каналом образуют непрерывный ряд и этот ряд совпадает с рядом их связывания с карбоксильной группой. Предполагают, что селективный фильтр кальциевого канала содержит карбоксильную группу. Все двух валентные катионы, которые связываются с этой группой слабее, чем кальций, проникают лучше (Ba+ и Sr+). Катионы, которые связываются с этой группой сильнее, чем кальций являются блокторами кальциевых каналов (Co2+,Ni2+,Cd2+).
Регулирующий центр кальциевого канала, как считают, располагается у устья, где есть связывающая структура, которая взаимодействует с двухвалентными катионами.
В бескальциевом растворе (с добавлением хелаторов Са2+ ЭГТА, ЭДТА) происходит модификация кальциевого канала: они сохраняют потенциал-зависимость, но теряют слективность и пропускают ионы натрия. По-видимому, в отсутствии ионов кальция устраняется селективный фильтр у устья канала. В нормальных условиях с этой структурой связан кальций, и в этих условиях канал сохраняет обычную избирательность по отношению к двухвалентным катионам. Если же ионы кальция удаляются из этого центра, то наступает конформационная перестройка канала, что ведет к потере его избирательности, и он начинает работать как обычный натриевый канал.
Модель Х1 иХ1 для натриевых и калиевых каналов применяются и к кальциевым каналам (перемещением заряженной частицы)
Проводимость (g) для кальциевого канала также зависит от активационной (m) и инактивацинной частиц (h):
gCa = gCa m2h (для натрия m3)
Ионный ток по закону Ома:
ICa =gCa m2h (E – ECa)
Если натриевый ток – быстрый входящий ток, то активация кальциевых каналов происходит медленнее, чем натриевых каналов. Кальциевый ток во многих электровозбудимых клетках недостаточно велик для того, чтобы самостоятельно (без натриевого тока) обеспечить регенеративный потенциал действия. Поэтому имеются мембраны, где именно кальциевые каналы обеспечивают входящий кальциевый ток, а передний фронт потенциала действия создает мощный натриевый ток, который обеспечивает деполяризацию мембрану. Эта деполяризация и активирует кальциевые каналы во многих клетках.
Процесс инактивации кальциевых каналов более сложный, чем натриевых каналов. У натриевых каналов происходит инактивация вследствие деполяризации мембраны, у кальциевых инактивация зависит от силы кальциевого тока. Чем выше ICa, тем быстрее инактивация, которая развивается в результате увеличения внутриклеточной концентрации ионов кальция [Ca2+]in. Если [Ca2+]in приблизительно достигает 10–7 – 10–6 М, то проводимость кальциевого канала обратимо подавлена.
Регуляция и модуляция активности кальциевых каналов может производиться многочисленными способами (факторами). На их активность могут влиять циклические нуклеотиды (цАМФ и цГМФ) и другие продукты метаболизма клетки.
Условно кальциевые каналы подразделяют на:
медленные:
1. порог активации – 30мВ
2. двухфазная инактивация
3. блокирование производными 1,4-дигидропиридина
быстрые:
1. порог активации – 70 – 60 мВ
2. быстрая инактивация
3. отсутствие чувствительности к 1,4дигидропиридинам
Дата добавления: 2015-06-22; просмотров: 2694;