Энергетический баланс аэробного распада глюкозы
1. При специфическом пути распада глюкозы образуется 2 молекулы пирувата, 2 АТФ (субстратное фосфорилирование) и 2 молекулы НАДН+Н+.
2. Окислительное декарбоксилирование каждой молекулы пирувата - 3 АТФ; 2 молекул пирувата дадут 6 молекул АТФ.
3. В результате сгорания ацетил-КоА в ЦТК и сопряженных ЦПЭ – 12 АТФ; 2 молекулы ацетил-КоА дадут 24 АТФ.
4. Малатный челночный механизм переносит НАДН+Н+ в митохондрии – 3 АТФ; 2 НАДН+Н+ дадут 6 АТФ.

Малатный челночный механизм
Итого: при распаде 1 молекулы глюкозы в аэробных условиях образуется 38 молекул АТФ. В некоторых тканях действует не малатный, а глицерофосфатный челночный механизм переноса восстановительных эквивалентов; в этом случае в митохондрии поступит 2 молекулы ФАДН2, которые в ЦПЭ дадут 4 молекулы АТФ и тогда общее количество при полном окислении глюкозы будет 36 АТФ.
По современным представлениям считается, что НАДН+Н+ в полной дыхательной цепи дает 2,5 АТФ, а ФАДН2 – 1,5 АТФ. Тогда полное окисление одной молекулы глюкозы будет сопровождаться выделением 32 (30) молекул АТФ (32 АТФ при малатном челночном переносе восстановительных эквивалентов через митохондриальную мембрану и 30 – при глицерофосфатном челночном механизме).
α-Глицерофосфатный челночный механизм переносит восстановительные эквиваленты за счет восстановления диоксиацетонфосфата до α-глицерофосфата с использованием НАДН+Н+ и фермента гицерофосфатдегидрогеназы. Эта реакция важна в жировой ткани при биосинтезе триацилглицеролов и глицерофосфолипидов.
В анаэробных условиях распад глюкозы до пирувата происходит аналогично, как и в аэробных условиях.
Судьба пирувата, образуемого при гликолизе, зависит от наличия или отсутствия кислорода. В анаэробных условиях в результате одиннадцатой реакции происходит восстановление пировиноградной кислоты и образуется молочная кислота (лактат). Реакция протекает при участии фермента лактатдегидрогеназы и кофермента НАДН+Н+, образованного при окислении 3-фосфоглицеролового альдегида в 1,3-бисфосфоглицероловую кислоту. Образование лактата позволяет регенерировать НАД+, который затем снова используется дегидрогеназой-3-фосфоглицеролового альдегида.

В гликолизе выделяют центральную окислительно-восстановительную реакцию гликолиза (гликолитическую оксидоредукцию), которая состоит в окислении 3-фосфоглицеролового альдегида с образованием НАДН+Н+ с последующим переносом атомов водорода от восстановленного НАД на пируват.
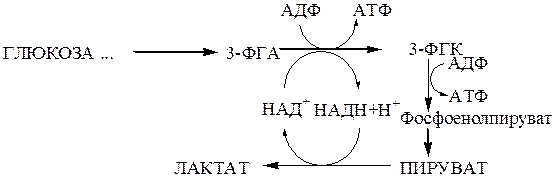
Гликолиз до лактата происходит только в анаэробных условиях(например, в мышцах при интенсивной мышечной работе) и в клетках,лишенных митохондрий (эритроциты).
Итак, энергетическая эффективность гликолиза в анаэробных условиях составляет 2 молекулы АТФ на одну молекулу глюкозы.
Анаэробному расщеплению могут подвергаться глюкозные остатки, отщепленные от концов цепей гликогена фосфоролитическим путем – гликогенолиз. Включение гликогена в процесс анаэробного распада осуществляется с помощью трех ферментов: 1) гликоген-фосфорилаза катализирует расщепление концевых α-1,4-гликозидных связей с выделением глюкозо-1-фосфата; 2) дебраншинг фермент катализирует расщепление α-1,6-гликозидных связей с выделением глюкозо-1-фосфата; 3) фосфоглюкомутаза катализирует превращение глюкозо-1-фосфата в глюкозо-6-фосфат. Поскольку при гликогенолизе на образование глюкозо-6-фосфата АТФ не тратится, в процессе гликогенолиза накапливаются не две, а три молекулы АТФ на глюкозный остаток. Однако это не означает, что гликогенолиз энергетически более выгоден, чем гликолиз, так как на синтез гликогена из глюкозы расходуется АТФ.
Итак, при расщеплении гликогена в анаэробных условиях (гликогенолиз) образуется 3 молекулы АТФ (не тратится АТФ на образование глюкозо-6-фосфата).
Эффект Пастера-ингибирование гликолиза (накопления лактата) кислородом. Механизмы эффекта:
1. При аэробном распаде углеводов не функционирует окислительно-восстановительная реакция гликолиза, поскольку восстановительные эквиваленты НАДН+Н+ из цитоплазмы переносятся в митохондрии. Поэтому НАДН+Н+ не может служить для восстановления пирувата в лактат.
2. При наличии аэробных условий образуются цитрат и АТФ, которые аллостерически тормозят активность фосфофруктокиназы (реакция лимитирует скорость всего процесса).
3. Дыхание подавляет гликолиз из-за конкуренции за АДФ между митохондриальной цепью переноса электронов и фосфоглицераткиназой и пируваткиназой гликолиза в цитоплазме. АДФ преимущественно используется в реакции окислительного фосфорилирования в митохондриях, а не субстратного фосфорилирования в гликолизе.
Дата добавления: 2015-06-12; просмотров: 10491;