Половое размножение
зачем нужен секс?
Почему и как появилось половое размножение – это одна из самых сложных тем в эволюционной биологии. Сотни идей и гипотез, аргументов и контраргументов. Основная проблема: затраты налицо, поэтому должен быть какой-то очень серьезный и быстрый выигрыш, иначе отбор не поддержит.
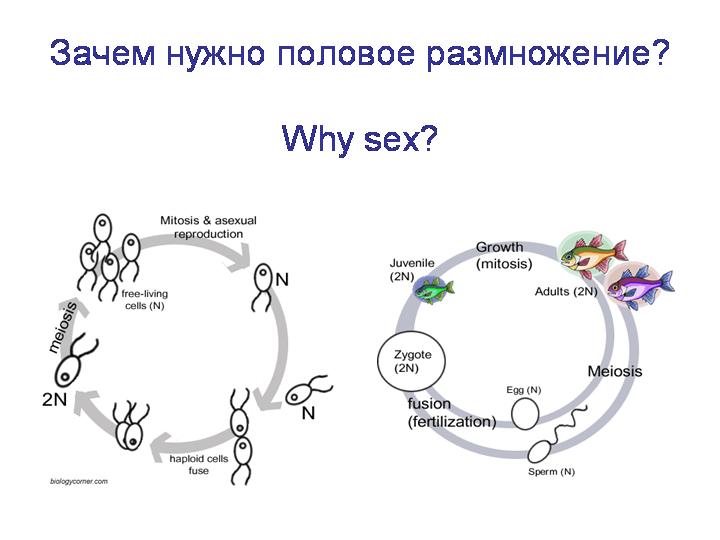
Гаплоидный и диплоидный жизненный цикл.
Начнем мы с того, что свяжем между собой половое размножение и другое явление, с которым мы уже знакомы – горизонтальный обмен генами. Как вы помните, у многоклеточных случаев горизонтального обмена генами известно мало. Однако в 2008 году было сделано потрясающее открытие, которое показало, что некоторые животные все-таки могут в массовом порядке заимствовать гены, причем не только у вирусов или МГЭ, а у клеточных организмов: у бактерий, грибов, растений.
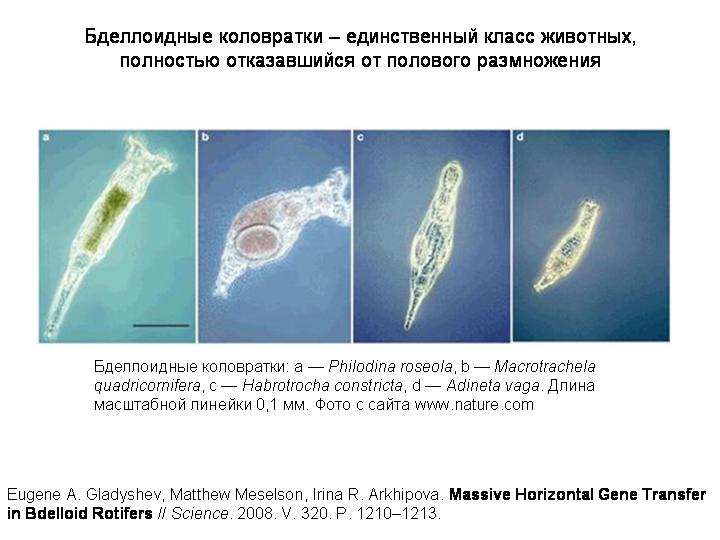
Эти животные – бделлоидные коловратки, группа во многом уникальная (...)
Самая удивительная особенность бделлоидных коловраток – полный отказ от полового размножения. Я уже говорил о возможных параллелях между горизонтальным генетическим обменом и половым размножением, и о том, что половое размножение в определенном смысле заменило эукариотам горизонтальный генетический обмен. И вот на примере бделлоидных коловраток мы видим подверждение этой идеи.
Животные вообще довольно легко утрачивают половое размножение и переходят к партеногенезу (...). Такое не раз происходило в разных эволюционных линиях. Однако виды животных, отказавшиеся от полового размножения, имеют тенденцию очень быстро вымирать. Они не успевают дивергировать (...) и дать начало, допустим, целому бесполому семейству или отряду. Современные бесполые виды, как правило, представляют собой концевые веточки на эволюционном дереве. Это значит, что те бесполые виды, которые появлялись раньше, как правило, быстро вымирали и не успевали дать начало другим бесполым видам.
Бделлоидные коловратки представляют собой удивительное исключение. Это целый класс животных (включающий около 400 видов), которые размножаются только бесполым путем (партеногенетически). Никто никогда не видел самцов бделлоидных коловраток. По-видимому, полового размножения не было уже у общего предка бделлоидных коловраток, который жил много десятков миллионов, возможно, около 100 миллионов лет назад.
Бделлоидные коловратки — главный камень преткновения для всех теоретиков, пытающихся объяснить биологический смысл полового размножения. Какое ни придумай объяснение, сразу же возникает «проклятый» вопрос: если половое размножение такое полезное, как же бделлоидные коловратки без него обходятся? И если бделлоидные коловратки научились без него обходиться, почему другие животные не пошли по тому же пути?
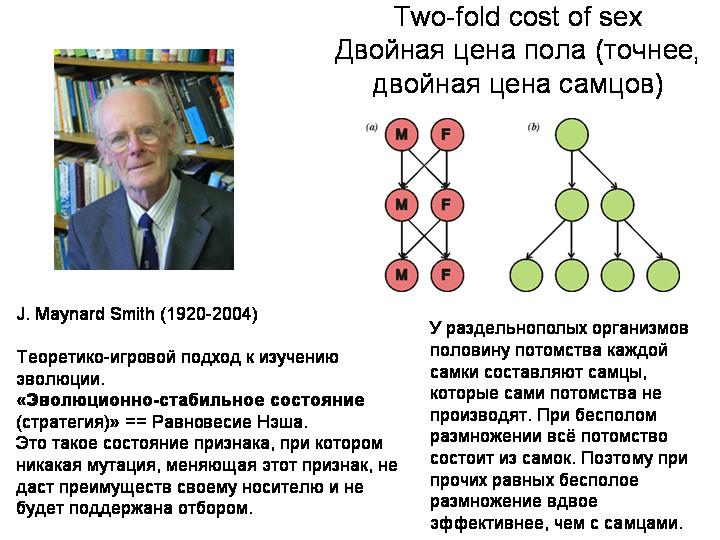
Ведь половое размножение — весьма «дорогое удовольствие» с точки зрения естественного отбора. При бесполом размножении вы передаете каждому потомку все свои гены, а при половом — только половину. За половое размножение приходится платить двукратным снижением эффективности передачи генов потомству.
Джон Мэйнард Смит назвал этот парадокс «двойной ценой пола». Здесь надо сделать небольшое отступление. Дж. Мэйнард Смит – один из величайших биологов-теоретиков 20 века. Одной из его заслуг является то, что он, вместе с некоторыми другими авторами, ввел в биологический обиход теорию игр и ввел такое понятие, как «эволюционно стабильное состояние» (или стратегия).
Эволюционно-стабильное состояние в биологии соответствует «равновесию Нэша» в теории игр. Равновесие Нэша — так в теории игр называется тип решений игры двух и более игроков, в котором ни один участник не может увеличить выигрыш, изменив своё решение (свою стратегию игры) в одностороннем порядке.
Соответственно, эволюционно-стабильное состояние – это такое состояние признака в популяции, при котором никакая мутация, меняющая этот признак, не даст преимуществ своему носителю и не будет поддержана отбором.
Много всего хорошего могло бы появиться в эволюции: например, всеобщая любовь и дружба между особями в каждой популяции, самопожертвование для общего блага и т.п. Но в большинстве случаев такие стратегии не являются эволюционно стабильными. Допустим, у нас есть вид животных, все особи которого друг за друга горой и готовы жертвовать жизнью ради спасения любых других представителей своего вида. Это было бы очень хорошо для вида в целом. Но эволюции нет дела до интересов вида. Такая ситуация не будет эволюционно стабильной, и вот почему.
Предположим, у нас есть такой вид, состоящий сплошь из героев-альтруистов. Что произойдет, если в ней появится мутант-эгоист? Он, к сожалению, почти наверняка получит адаптивное преимущество. Он будет пользоваться добротой других особей, а сам ничего не будет давать взамен. В общем случае его стратегия будет более выигрышной – он оставит больше потомства. Соответственно «ген эгоистичного поведения» будет распространяться! В итоге ген эгоизма может полностью вытеснить ген альтруизма. Иногда, при особых условиях, этого не происходит, но все-таки чаще, как правило, в природе живые существа ведут себя эгоистично, хоть это и плохо для вида. Это плохо для вида, но выгодно для самих эгоистичных особей и для их генов, которые благодаря этому эгоизму распространяются лучше, чем гены альтруистов.
У нас будет специальная тема «эволюция альтруизма», и мы подробно разберем ситуации, при которых альтруизм все-таки развивается. Но в общем случае, как правило, если нет каких-то особых условий, альтруизм – эволюционно нестабильная стратегия. Она вытесняется мутантами-эгоистами.
Вернемся к половому размножению. Половое размножение — «дорогое удовольствие» с точки зрения естественного отбора. Джон Мейнард Смит назвал центральную проблему, с которой сталкиваются двуполые виды – имеющие самцов и самок – проблемой «двойной цены пола». Суть идеи в том, что разделение на самцов и самок должно быть эволюционно нестабильным.
У раздельнополых организмов половину потомства каждой самки составляют самцы, которые сами потомства не производят. При бесполом размножении всё потомство состоит из самок. Поэтому при прочих равных бесполое размножение вдвое эффективнее, чем с самцами.
Следовательно, раздельнополость должна быть эволюционно нестабильна. Это не равновесие по Нэшу. Представьте, что у нас есть популяция двуполых животных, и в ней вдруг появляется мутантная самка, способная размножаться партеногенезом.
Потомство этой самки, очевидно, должно очень быстро вытеснить потомство всех остальных самок. Просто потому, что партеногенетические самки будут размножаться вдвое быстрее, чем самки, вынужденные спариваться с самцами и рожать не только дочерей, но и сыновей.
Предполагается, что каждая самка может произвести на свет ограниченное количество потомков – будь то сыновья или дочери (например, только двух, как на рисунке). Если она производит только дочерей, способных обходиться без самцов, ее потомство будет размножаться ровно вдвое быстрее.
В результате гены раздельнополых особей будут вытеснены генами партеногенетических особей. Это и значит, что раздельнополость эволюционно нестабильна.
Из этого, далее, следует, что если, несмотря на двойную цену самцов у многих организмов все-таки есть самцы, то это должно давать какое-то преимущество. Настолько значительное, что оно перевешивает эту двойную цену.
----------------------------------------------------
Теперь вернемся к бделлоидным коловраткам. Они отказались от полового размножения, что вроде бы соответствует идеям Мэйнарда Смита. Отказались, потому что это позволяет им не платить «двойную цену». Но, с другой стороны, они при этом должны были потерять и то преимущество, в чем бы оно ни заключалось, которое у других животных перевешивает двойную цену самцов. Многие авторы отмечали, что любое теоретическое построение, указывающее на преимущества полового размножения, обязательно должно как-то объяснять парадокс бделлоидных коловраток.
И вот, похоже, удалось нащупать решение этой давней загадки. В хромосомах бделлоидной коловратки обнаружено множество генов, не встречающихся ни у каких других животных. Некоторые из этих генов явно имеют бактериальное происхождение. Другие происходят от грибов, третьи — от растений. Были выявлены десятки генов, заимствованных коловратками у представителей других царств.
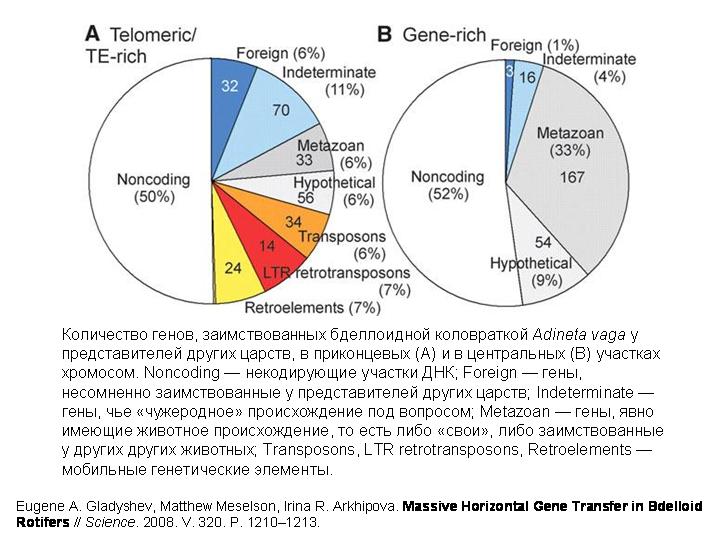
Некоторые заимствованные бактериальные гены у коловраток сохранили структуру, характерную для прокариотических генов (в них нет интронов), а другие уже успели обзавестись интронами, характерными для эукариот (как – неизвестно). По крайней мере некоторые из заимствованных генов реально работают в клетках коловраток и кодируют функциональные белки.
Примененные методы не позволяли выявить гены, заимствованные у других животных или, тем более, у других бделлоидных коловраток. Выявлялись только гены, заимствованные не у животных.
Животные стараются оберегать свои половые клетки от чужого генетического материала, в том числе вирусного. У бделлоидных коловраток эти барьеры, по-видимому, ослаблены. Это может быть связано с необычным образом жизни этих животных. Они живут в мелких лужах и отлично переносят высыхание. Потом их, как пыль, может перенести ветром в другую лужу. Однако при высыхании мембраны клеток могут повреждаться, что облегчает проникновение чужой ДНК. При высыхании образуются разрывы в хромосомах, которые клеткам приходится зашивать, репарировать, когда коловратка снова размокнет. В ходе репарации разорванных хромосом имеется вероятность случайного включения в хромосому чужеродного фрагмента.
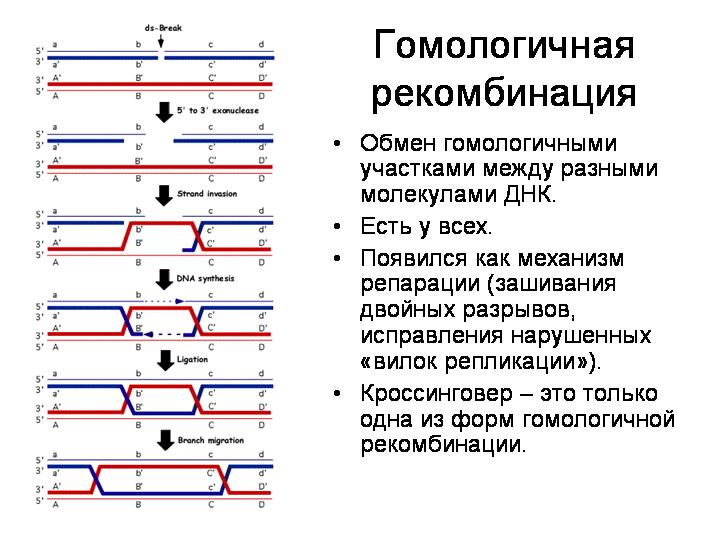
При высыхании образуются разрывы в ДНК, которые клеткам приходится репарировать при помощи гомологичной рекомбинации. При этом есть вероятность включения в хромосому чужеродного фрагмента.
Пока еще не известно, обмениваются ли бделлоидные коловратки генами между собой. Однако вряд ли коловратки, охотно заимствующие гены у микробов и растений, имеют при этом какую-то особую систему защиты от заимствования генов у родственников. Если же они хотя бы иногда меняются генами друг с другом, то получается, что они на самом деле не отказались от межорганизменной генетической рекомбинации, то есть перемешивания генов разных родителей в геноме потомства. Они просто вернулись от продвинутого варианта такой рекомбинации (полового процесса) к более примитивному варианту — горизонтальному обмену.

Этот пример подтверждает, что половой процесс и горизонтальный обмен генами – явления родственные. Судите сами: бделлоидные коловратки – единственная группа животных, которая давно отказалась от полового размножения и до сих пор не вымерла. С другой стороны, единственная группа животных, у которой обнаружен массовый горизонтальный обмен генами – это опять-таки бделлоидные коловратки. Такое совпадение не может быть случайным.
В чем же состоит это загадочное преимущество, которое дает половой процесс? Преимущество, утратив которое, виды, как правило, быстро вымирают. Преимущество, из-за которого большинство животных все-таки не уподобились бделлоидным коловраткам. Которое должно проявляться быстро, а не через миллион лет.

Рассмотрим две главные гипотезы. Первая известна под названием «храповик Мёллера». Храповик — это устройство, в котором ось может крутиться только в одну сторону. Суть в том, что если у бесполого организма возникает вредная мутация, его потомки уже не могут от нее избавиться. Она будет, как родовое проклятие, передаваться всем его потомкам вечно (если только не произойдет обратная мутация, а вероятность этого очень мала).
У бесполых организмов отбор может отбраковывать только целые геномы, но не отдельные гены. Если в бесполой популяции случайно погибает особь с наименьшим числом вредных мутаций, это означает необратимое ухудшение, необратимую потерю. При отсутствии рекомбинации (обмена гомологичными генами между особями) невозможно получить из сильно отягощенных мутациями геномов менее отягощенный.
Поэтому в череде поколений бесполых организмов может (при соблюдении определенных условий) происходить неуклонное накопление вредных мутаций. Одним из таких условий является большой размер генома. Чем больше в геноме генов, тем больше новых мутаций будет в среднем у каждого организма. Поэтому бесполые организмы не могут позволить себе иметь большие геномы – точнее, большие осмысленные части геномов.
Бесполые организмы не могут себе позволить иметь маленькие популяции (и крупные размеры), малое число потомков (т.к. только очень эффективный очищающий отбор может их спасти).
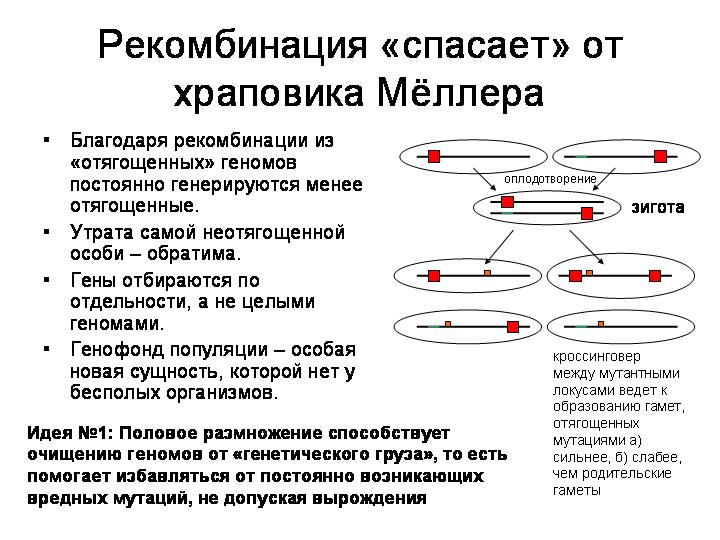
Если же организмы размножаются половым путем, то индивидуальные геномы постоянно рассыпаются и перемешиваются (в результате мейозов и слияний гамет), а новые геномы формируются из фрагментов, ранее принадлежавших разным организмам. В результате возникает особая новая сущность, которой нет у бесполых организмов, — генофонд популяции. Гены получают возможность размножаться или выбраковываться независимо друг от друга. Ген с неудачной мутацией может быть отбракован отбором, а остальные («хорошие») гены данного родительского организма могут благополучно сохраниться в популяции.
Таким образом, первая идея состоит в том, что половое размножение способствует очищению геномов от «генетического груза», то есть помогает избавляться от постоянно возникающих вредных мутаций, не допуская вырождения (снижения общей приспособленности популяции).
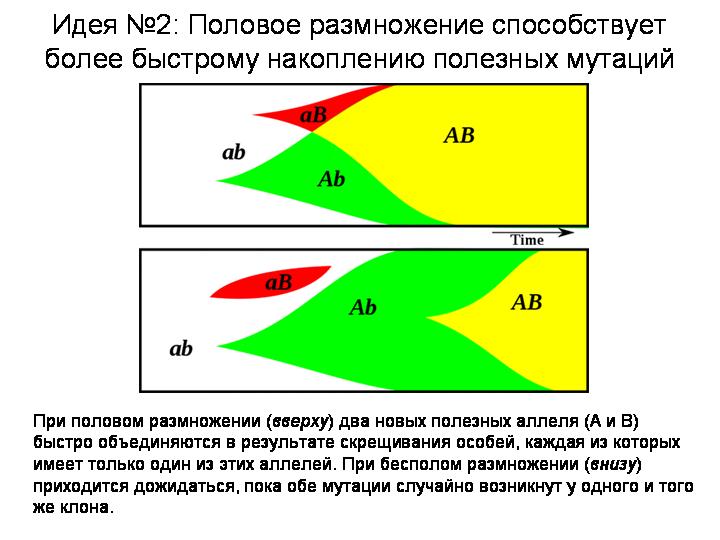
Вторая идея: половое размножение помогает эффективнее адаптироваться к меняющимся условиям за счет ускоренного накопления мутаций, полезных в данной обстановке. Допустим, у одной особи возникла одна полезная мутация, у другой — другая. Если эти организмы бесполые, у них мало шансов дождаться объединения обеих мутаций в одном геноме. Половое размножение дает такую возможность. Оно фактически делает все полезные мутации «общим достоянием». Ясно, что скорость приспособления к меняющимся условиям у организмов с половым размножением должна быть выше.
Все эти теоретические построения, однако, основаны на определенных допущениях. Результаты математического моделирования свидетельствуют о том, что степень полезности или вредности полового размножения по сравнению с бесполым размножением или самооплодотворением зависит от многих параметров. В их числе размер популяции; скорость мутирования; размер генома; количественное распределение мутаций в зависимости от степени их вредности/полезности; число потомков, производимых одной самкой; эффективность отбора (степень зависимости числа оставляемых потомков не от случайных, а от генетических факторов) и т. д. Некоторые из этих параметров очень трудно измерить не только в природных, но и в лабораторных популяциях.

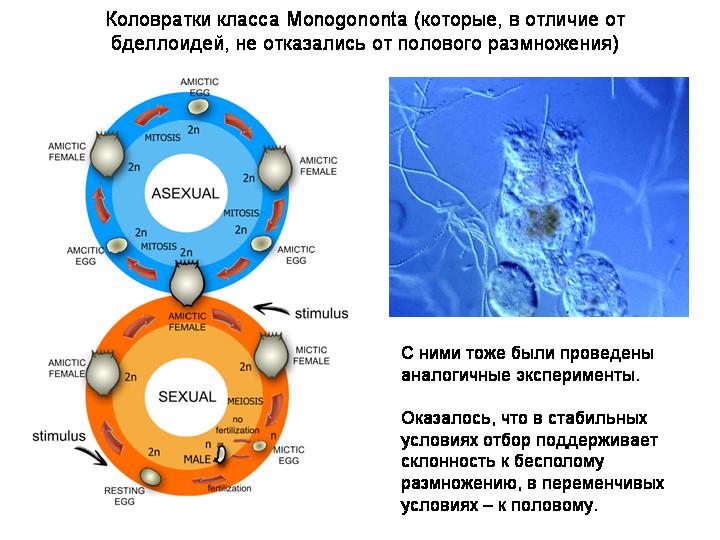
Поэтому гипотезы остро нуждаются в экспериментальной проверке. Такие проверки были проведены. Рассмотрим одно исследование, выполненное на круглом черве Caenorhabditis elegans (биологи из университета штата Орегон, 2009 г). Это исследование показало действенность обоих механизмов, обеспечивающих преимущество тем популяциям, которые не отказываются от самцов, несмотря на их «двойную цену».
Уникальный объект для изучения роли самцов. Черви Caenorhabditis elegans словно нарочно созданы для проверки таких гипотез.
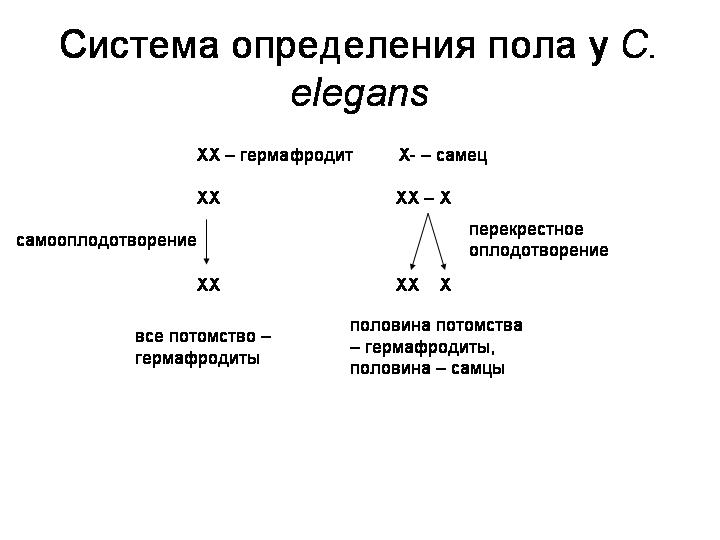
У этих червей нет самок. Популяции состоят из самцов и гермафродитов, причем последние преобладают. У гермафродитов две X-хромосомы, у самцов — только одна (система определения пола X0). Гермафродиты производят сперматозоиды и яйцеклетки и могут размножаться без посторонней помощи путем самооплодотворения. Самцы производят только сперматозоиды и могут оплодотворять гермафродитов. В результате самооплодотворения на свет появляются только гермафродиты. При перекрестном оплодотворении половина потомства оказывается гермафродитами, половина — самцами.
Обычно частота перекрестного оплодотворения в популяциях C. elegans не превышает нескольких процентов. Чтобы определить эту частоту, не обязательно наблюдать за интимной жизнью червей — достаточно знать процент самцов в популяции.
Следует пояснить, что самооплодотворение — это не совсем то же самое, что бесполое (клональное) размножение, однако различия между ними не очень существенны (они быстро сходят на нет в череде самооплодотворяющихся поколений. Самооплодотворяющиеся организмы за несколько поколений неизбежно становятся гомозиготными по всем локусам. После этого потомство перестает отличаться от родителей генетически, точно так же, как и при клональном размножении.
У C. elegans известны мутации, влияющие на частоту перекрестного оплодотворения. Одна из них смертельна для самцов и фактически приводит к тому, что в популяции остаются только гермафродиты, размножающиеся путем самооплодотворения. Другая мутация лишает гермафродитов способности производить сперму и фактически превращает их в самок. Популяция, в которой все особи несут эту мутацию, становится обычной раздельнополой популяцией, как у большинства животных.
Авторы сравнивали три породы червей с одинаковыми геномами, различающимися только наличием этих мутаций.
· «Дикий тип» (контроль). Частота перекрестного оплодотворения около 5%.
· Мутанты с облигатным самооплодотворением (самцов нет).
· Мутанты с облигатным перекрестным оплодотворением (обычная раздельнополая популяция).
С этими породами провели два эксперимента.
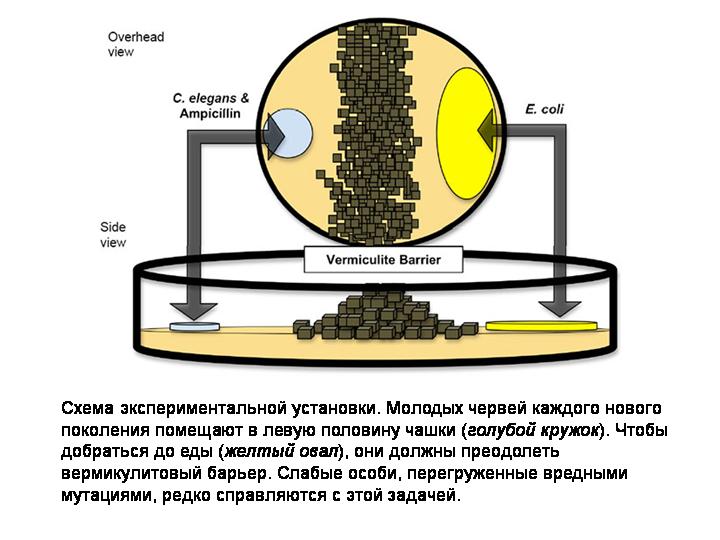
В первом эксперименте проверялась гипотеза о том, что перекрестное оплодотворение помогает избавляться от «генетического груза». Эксперимент продолжался в течение 50 поколений (червей, а не экспериментаторов). Червей подвергали действию химического мутагена — этилметансульфоната. Это приводило к увеличению частоты мутирования в четыре раза. Молодых животных помещали в чашку Петри, разделенную пополам труднопреодолимой стенкой (см. на слайде), причем червей сажали в одну половину чашки, а их пища — бактерии E. coli — находилась в другой половине. Чтобы добраться до еды, а значит получить шанс выжить и оставить потомство, черви должны были преодолеть препятствие. Тем самым экспериментаторы повысили эффективность очищающего отбора, который отсеивает вредные мутации. В обычных лабораторных условиях эффективность отбора низкая, потому что черви окружены пищей со всех сторон. В такой ситуации могут выжить и размножиться даже очень слабые, перегруженные вредными мутациями животные. В новой экспериментальной установке этой уравниловке был положен конец. Чтобы переползти через стенку, червь должен быть здоров и крепок.
Авторы сравнивали приспособленность у червей до и после эксперимента, то есть у особей первого и пятидесятого поколения. Червей C. elegans можно хранить в замороженном виде. Это облегчает подобные эксперименты. Пока длился эксперимент, выборка червей 1-го поколения спокойно лежала в морозильнике. Приспособленность измеряли так. Червей смешивали в равной пропорции с контрольными червями, в геном которых был вставлен ген светящегося белка, и сажали в экспериментальную установку. Животным давали время, чтобы преодолеть барьер и размножиться, а затем определяли процент несветящихся особей в потомстве. Если этот процент увеличился в пятидесятом поколении по сравнению с первым — значит, в ходе эксперимента приспособленность выросла, если уменьшился — значит, имело место вырождение.
Результаты получились следующие. Подтвердилось предположение, что перекрестное оплодотворение помогает бороться с генетическим грузом. Чем выше частота перекрестного оплодотворения, тем лучше был итоговый результат (тем меньше выродились черви). Искусственно повышенная скорость мутирования привела к вырождению (снижению приспособленности) всех пород червей, кроме «облигатных перекрестников».
Самое интересное, что частота перекрестного оплодотворения у «диких» червей в ходе эксперимента стала заметно выше исходных 5%. Это означает, что в жестких условиях (имеется в виду как необходимость перелезать через барьер, так и повышенный темп мутагенеза) естественный отбор дает преимущество особям, размножающимся путем перекрестного оплодотворения. Потомство таких особей оказывается более жизнеспособным, и поэтому в ходе эксперимента происходит отбор на склонность к перекрестному оплодотворению.
Таким образом, первый эксперимент подтвердил гипотезу о том, что перекрестное оплодотворение помогает избавляться от вредных мутаций.

Во втором эксперименте проверялось, помогает ли перекрестное оплодотворение вырабатывать новые адаптации путем накопления полезных мутаций. На этот раз червям, чтобы добраться до пищи, нужно было преодолеть зону, заселенную патогенными бактериями Serratia. Эти бактерии, попадая в пищеварительный тракт C. elegans, вызывают у червя опасное заболевание. Чтобы выжить в этой ситуации, черви должны были либо научиться не глотать вредных бактерий, либо выработать устойчивость к ним. Какой из вариантов выбрали подопытные популяции — неизвестно, однако за 40 поколений черви облигатные перекрестники отлично приспособились к новым условиям, дикие черви приспособились хуже, а облигатные самооплодотворители не приспособились совсем (их выживаемость в среде с вредными бактериями осталась на исходном низком уровне). И снова в ходе эксперимента у диких червей под действием отбора возросла частота перекрестного оплодотворения.
Таким образом, перекрестное оплодотворение действительно помогает популяции приспосабливаться к новым условиям, в данном случае — к появлению болезнетворного микроба. Тот факт, что в ходе эксперимента у диких червей увеличивалась частота перекрестного оплодотворения, означает, что спаривание с самцами дает гермафродитам немедленное адаптивное преимущество, которое, очевидно, перевешивает «двойную цену», которую им приходится платить, производя на свет самцов.
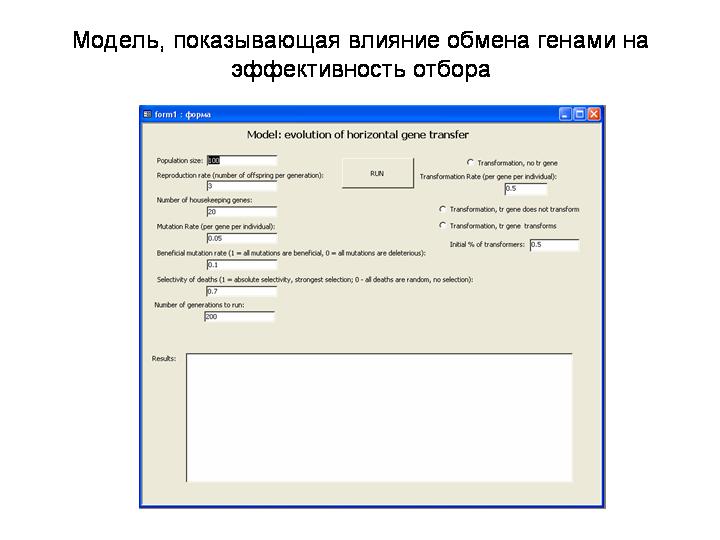
Модель эволюции горизонтального генетического обмена (на примере естественной трансформации)
Моделируется популяция гаплоидных организмов, размножающихся бесполым путем. 100, 3, 20, 0.05, 0.1, 0.7, 200
Параметры модели:
Population size – размер популяции (количество взрослых организмов в каждом поколении),
Reproduction rate – число потомков, производимых каждым взрослым организмом в каждом поколении (произведя потомков, взрослый организм погибает),
Number of housekeeping genes – Число «генов» (или генных комплексов), соответствующих каким-либо жизнеобеспечивающим функциям организма. У каждого гена каждого организма есть величина fitness (F, приспособленность), показывающая, с какой эффективностью данный ген выполняет свою функцию. Изначально все особи в популяции получают значения F=100 для каждого гена.
Mutation rate – частота мутирования (на 1 ген). Мутации возникают при образовании потомков. Потомок получает гены родительской особи, причем с вероятностью mutation rate каждый ген может мутировать. Мутация делятся на полезные и вредные. Полезные мутации увеличивают fitness гена на случайное число в диапазоне от 0 до 1, вредные – снижают fitness гена на случайное число в диапазоне от 0 до 5.
Beneficial mutation rate (BMR) – доля полезных мутаций (например, при Beneficial mutation rate = 0.1 каждая мутация с вероятностью 0.1 будет полезной, с вероятностью 0.9 – вредной). Таким образом, при заданных здесь параметрах «средний вред» мутаций превышает их «среднюю пользу» в 45 раз (вредные мутации возникают в 9 раз чаще и приносят в среднем в 5 раз больше вреда, чем полезные – пользы).
Selectivity of deaths (SD) – степень зависимости смертности потомства от генов, а не от случая. Например, при Selectivity of deaths = 0.7 гибель молодой особи на 70% определяется суммарной приспособленностью ее генов, на 30% - случайностью.
Все молодые особи, родившиеся в данном поколении, ранжируются в порядке убывания величины SD * (Fi - MinFit) / (MaxFit - MinFit) + Rnd * (1 - SD), где Fi – суммарная F всех геннов данной (i-ой) особи, MinFit – минимальная, а MaxFit – максимальная приспособленность молодых особей в данном поколении. Первые PS особей из данного списка становятся взрослыми, остальные погибают. Тем самым реализуется зависимость смертности от сочетания генетических и случайных причин, причем вклад генетических причин равен SD.
Дата добавления: 2015-06-10; просмотров: 2733;