Особенности прорастания семян
Сформировавшиеся семена подавляющего большинства цветковых растений состоят из зародыша, эндосперма и семенной кожуры. Однако соотношение зародыша и эндосперма сильно варьирует. В одних случаях зародыш мал и ткань эндосперма занимает большую часть объема (злаки, лилейные, лютиковые). В других зародыш ко времени созревания сильно разрастается и от эндосперма остается лишь небольшой слой клеток под семенной кожурой (яблоня, тыква и др.) или ничего не остается, а зрелое семя состоит из зародыша и семенной кожуры (многие бобовые). В некоторых случаях помимо эндосперма в семенах имеется ткань, также заполненная питательными веществами, но развивающимися не из триплоидного ядра зародышевого мешка, а из нуцеллуса (перисперм).
В соответствии с преимущественным содержанием тех или иных запасных питательных веществ семена делят на масличные, крахмалистые и белковые. В качестве примера масличных можно привести семена клещевины, содержание жира в которых достигает 64% на сухое вещество. Наивысшее содержание крахмала характерно для семян злаков. Так, семена кукурузы содержат до 72% крахмала. Белки содержатся во всех семенах. Они откладываются в виде алейроновых зерен, эластичных волокон или бесформенной массы. Высокое содержание белков характерно для бобовых растений. Так, семена сои содержат до 40% белка.
Семена всех растений содержат фитин. Главная функция фитина — снабжать зародыш соединениями фосфора. Одновременно фитин содержит некоторое количество К, Mg, Ca. В семенах содержатся также ферменты и гормоны, однако в неактивном состоянии. Распределение веществ в семенах неравномерно. Ткани зародыша обогащены минеральными элементами. Так, содержание золы в зародыше составляет 5—5,5%, тогда как в эндосперме—всего около 0,5%. Показано, что зародыш обогащен активными веществами (ферменты, аминокислоты, гормоны), причем особенно много их в зародышевом корешке. Последнее как бы уже предопределяет ту специфическую роль, которую играют клетки корня в жизнедеятельности растущего организма.
Процесс прорастания семян включает в себя и те процессы, которые происходят в семени до того, как появляются признаки видимого роста. Для прорастания необходимы определенные условия. Прежде всего нужна вода. Воздушно-сухие семена содержат до 20% воды и находятся в состоянии вынужденного покоя. Сухие семена быстро поглощают воду и набухают. Набухание обратимо: если еще не началось деление и растяжение клеток зародыша, то семена можно подсушить и они не потеряют жизнеспособности. Для процесса прорастания необходим кислород, поддерживающий процесс дыхания, поскольку процессы, происходящие на первых этапах прорастания, идут с затратой энергии. Оптимальные температуры для прорастания семян обычно соответствуют тем, которые характерны для ареала распространения данного вида растений. Семена некоторых растений лучше прорастают при сменной температуре. Прорастание семян ряда растений требует выдерживания при пониженных температурах. Есть растения, для прорастания семян которых необходим свет.
Вода поступает в семена по градиенту водного потенциала, согласно уравнению: Ψв = Ψосм + Ψдавл. + Ψматр. Согласно развиваемым представлениям (НА. Аскоченская) поступление воды в семена можно разделить на три этапа. Первый этапосуществляется в основном за счет матричного потенциала, или сил гидратации. Гидратация — спонтанный процесс. Находящиеся в семени запасные питательные вещества содержат большое количество гидрофильных группировок, таких, как —ОН, —СООН, —NH2. Молекулы воды вокруг гидра-тированных веществ принимают льдоподобную структуру. Притягивая молекулы воды, гидрофильные группировки уменьшают ее активность. Водный потенциал становится более отрицательным, вода устремляется в семена. На втором этапепоглощения воды силы набухания, или матричный потенциал, также являются основными. Однако начинают играть роль осмотические силы — осмотический потенциал, поскольку в этот период происходит интенсивный гидролиз сложных соединений на более простые. На третьем этапе,который наступает в период наклевывания семян, когда клетки растягиваются и появляются вакуоли, главной силой, вызывающей поступление воды, становятся осмотические силы — осмотический потенциал. Уже в процессе набухания семян начинается мобилизация питательных веществ — жиров, белков и полисахаридов. Это все нерастворимые, плохо передвигающиеся сложные органические вещества. В процессе прорастания происходит перевод их в растворимые соединения, легко используемые для питания зародыша, поэтому необходимы соответствующие ферменты.
Частично ферменты находятся в эндосперме или зародыше в связанном, неактивном состоянии и под влиянием набухания переходят в активное состояние. Однако ферменты образуются и de novo. Для образования ферментов de novo необходимы соответствующие матричные РНК. Согласно современным представлениям (Дюр) матричную РНК в прорастающих семенах можно разделить на три типа (по времени образования). К первому типу можно отнести так называемую предсуществующую (предобразованную — остаточную) мРНК, которая была транскрибирована с ДНК еще в период эмбриогенеза семян. Эта РНК использовалась в эмбриогенезе для синтеза белков и теперь в процессе прорастания вновь может быть использована. Однако ее немного и ее роль невелика. Ко второму типу относится мРНК, также транскрибированная в эмбриогенезе, однако не прошедшая процессинга и поэтому неактивная. В процессе набухания она проходит все необходимые превращения (процессинг) и обеспечивает синтез белков, специфичных для прорастания, главным образом ферментов гидролиза. Третий тип — это новообразованная РНК, которая появляется через 1—2 ч после намачивания. Эта РНК транскрибируется с ДНК в процессе прорастания РНК-полимеразой-1 и также ответственна за синтез специфических белков-ферментов. Существуют данные, что в синтезе белка при прорастании сначала участвуют рибосомы, образованные еще в эмбриогенезе, затем, начиная примерно с 8 ч от намачивания семян, происходит усиленное образование рибосомальной РНК и формируются новые рибосомы.
Крахмал распадается до Сахаров в основном под влиянием двух ферментов — α- и β-амилазы. Показано, что (β-амилаза находится в сухих семенах в связанном (неактивном) состоянии, а α -амилаза вновь образуется в процессе прорастания. Одновременно происходит новообразование ферментов, катализирующих распад белков — протеаз, нуклеиновых кислот — нуклеаз. Вновь образуются также ферменты, участвующие в превращении жиров,— изоцитратлиаза и малатсин-тетаза. Большую роль в.регуляции образования ферментов играют фитогормоны, в частности гиббереллины. На семенах ячменя было показано, что если удалить зародыш, то ряд ферментов не образуется. Если эндосперм, лишенный зародыша, обработать гормоном гиббереллином, то в алейроновом слое появляются α -амилаза и протеазы. При этом установлено, что если эндосперм замачивать в тяжелой воде Н2О, то α -амилаза содержит 18О. Следовательно, под влиянием гиббереллина он синтезируется заново. Источником гиббереллина в семени является зародыш. В сухих семенах гиббереллины находятся в связанном состоянии. Они активизируются и частично образуются вновь под влиянием поступившей воды. Значение гиббереллина для индукции новообразования ферментов в процессе прорастания показано в основном на семенах однодольных растений. По-видимому, у разных растений в этом процессе могут участвовать и другие фитогормоны.
Под воздействием соответствующих ферментов начинается усиленная мобилизация — распад сложных нерастворимых соединений на простые растворимые. Так, крахмал распадается на сахара, белки расщепляются до аминокислот, а последние до органических кислот и аммиака. Образовавшийся аммиак обезвреживается в виде амидов. В процессах прямого аминирования и переамини-рования образуются новые аминокислоты, которые перетекают в осевую часть зародыша, где из них строятся новые специфические белки. Специфичность этих белков закодирована в ДНК клеток зародыша.
Более сложные превращения претерпевают жиры. Жиры сравнительно с углеводами — восстановленные соединения, и при их окислении выделяется больше энергии. Возможно, поэтому растения с мелкими семенами в большинстве случаев накапливают жиры как запасное питательное вещество. Распад жиров начинается с образования жирных кислот и глицерина. Процесс катализируется двумя липазами, одна из которых находится непосредственно в олеосомах, а другая в мембране глиоксисом — специальных органелл.
Жирные кислоты подвергаются в глиоксисомах окислению с образованием ацетил-КоА. Ацетил-КоА вступает в глиоксилатный цикл (рисунок 1). Прежде всего, при взаимодействии ацетил-КоА с щавелево-уксусной кислотой (ЩУК) образуется лимонная кислота. Лимонная кислота изомеризуется до изолимонной.
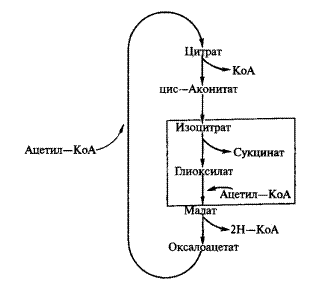
Рисунок 1 – Глиоксилатный цикл
Изолимонная кислота под влиянием уникального фермента этого цикла — изоцитратлиазы распадается на янтарную и глиоксилевую кислоты. В глиоксисомах оставшаяся глиоксилевая кислота со второй молекулой ацетил-КоА образует малат. Образовавшийся малат окисляется до ЩУК, и, таким образом, глиоксилатный цикл заканчивается. Янтарная кислота поступает в митохондрии, где превращается в реакциях цикла Кребса последовательно в фумаровую и ЩУК. ЩУК превращается в фосфоенолпировиноградную (ФЕП). Реакция катализируется ферментом ФЕП-карбоксилазой, требует затраты энергии. ФЕП в ряде реакций, обратных гликолизу, превращается в глюкозофосфат и далее в сахарозу. Глицерин превращается в дегидрооксиацетон и далее также в сахарозу. Образовавшиеся в результате распада растворимые соединения либо используются как дыхательный материал, либо транспортируются в зародыш. Таким образом, при прорастании семян идут многочисленные взаимосвязанные процессы — распад питательных веществ, их превращение, транспорт и образование из них новых веществ, идущих на построение клеток и органов. Энергия для всех этих процессов поставляется процессом дыхания.
Уже в первый период после намачивания интенсивность поглощения кислорода возрастает. Особенно сильно увеличивается интенсивность гексозомо-нофосфатного пути, а затем гликолиза. Возрастание интенсивности дыхания сопровождается усилением накопления АТФ, что и является, в свою очередь, необходимым условием для процессов обмена. Через 10—12 ч от начала набухания митохондрии усиленно растут и дифференцируются. Часть митохондрии, деградировавшая в период созревания семян, реактивируется. В дальнейшем, через 24 ч, происходит деление митохондрии, их число резко возрастает. Процесс окислительного фосфорилирования резко интенсифицируется и становится основным источником образования АТФ. Образовавшиеся в процессе распада соединения перетекают в осевую часть зародыша и там используются частично как дыхательный материал, а частично для построения веществ, необходимых для роста новых клеток и органов. Так, поступившие в осевую часть зародыша питательные вещества расходуются на построение новых белков и нуклеиновых кислот, компонентов клеточных оболочек (целлюлозы, пектиновых веществ), а также различных липидов, входящих в состав мембран.
Наконец, необходимо сказать, что на этой фазе прорастания образуются и такие фитогормоны, как ауксины и цитокинины. Согласно гипотезе, предложенной Овербиком, последовательность образования фитогормонов при прорастании идет следующим путем. Как уже рассматривалось выше, гиббереллины при набухании переходят в свободную форму и вызывают новообразование ферментов гидролаз в клетках алейронового слоя. Образовавшиеся при этом нуклеазы катализируют распад нуклеиновых кислот, появляются пуриновые основания, дают начало фитогормону цитокинину. Одновременно под влиянием ферментов протеаз белки распадаются до аминокислот, среди которых имеется триптофан. Как известно, триптофан является предшественником фитогормона ауксина.
Цитокинины и ауксины регулируют рост зародыша (эмбриогенез). Цитокинины вызывают деление клеток, ауксины — их растяжение. Росту органов зародыша способствуют такие ферменты, как пектинаэа и целлюлаза. Эндораманназы расщепляют маннаны (полисахариды), соединяющие фибриллы целлюлозы, и тем самым облегчают преодоление зародышевой осью сопротивления эндосперма. АБК ингибирует образование фермента манназы, и это одна из причин тормозящего влияния этого фитогормона на прорастание семян.
Увеличение длины осевых органов зародыша начинается раньше видимого процесса проклевывания. Запуск ростовых процессов осуществляется, в первую очередь, за счет растяжения клеток и начинается, когда матричный потенциал близок к нулю. Для того чтобы началось деление ядра (митоз), необходима редупликация ДНК. Показано, что синтез ДНК в прорастающих семенах начинается значительно позже синтеза РНК и белков. В сухих семенах основная часть клеток находится в состоянии g1 (в предсинтетическом периоде). Перед началом деления клетки переходят в период синтетический, после чего наступает период g2 (постсинтетический). Митозы происходят не во всех клетках зародыша одновременно. По-видимому, прежде всего к митозу переходят клетки зародышевого корешка.
Зародыш состоит из зародышевого корешка, стебля и почечки. Дальнейший рост проростков несколько различается в зависимости от типа растений. У однодольных растений проклевывание начинается с роста зародышевого корешка, затем при подземном прорастании начинает расти колеоптиль (первый свернутый лист). Когда колеоптиль достигает поверхности почвы, его рост прекращается, из почечки начинает расти первый настоящий лист, который прорывает колеоптиль. У многих однодольных растений из нижней части стебля образуются придаточные корни. У однодольных при надземном прорастании (лук) после появления корешка начинается неравномерное разрастание семядоли, которая, образуя петлю, выходит на поверхность.
У двудольных растений, если семядоли выносятся на поверхность (надземное прорастание), прежде всего вытягивается гипокотиль — подсемядольное колено. Гипокотиль растет неравномерно и образует петлеобразный изгиб, что позволяет лучше преодолевать сопротивление почвы. Имеются данные, что образование крючка гипокотиля связано с фитогормоном этиленом. Под влиянием света синтез этилена тормозятся, гипокотиль выпрямляется и выносит семядоли на поверхность. После этого трогается в рост почечка, которая дает начало главному стеблю, и формируются надземные побеги. При подземном прорастании семядоли на поверхность не выносятся, в этом случае вытягивается эпикотиль — надсемядольное колено. При этом на поверхность выносится хорошо сформированная почечка. Последовательность роста частей зародыша выработалась в процессе эволюции, что позволяет в первый период прорастания обеспечить прикрепление растения к субстрату и снабжение его водой.
Для нормального роста и формообразовательных процессов проростка необходим свет. В течение первых фаз прорастания проросток питается за счет готовых органических веществ, находящихся в эндосперме или семядолях. Как только появляются первые зеленые листья, начинается фотосинтез. На этом фаза проростка заканчивается, и растение переходит в ювенильную фазу. На свету в клетках листьев из протопластид развиваются пластиды. Еще до завершения роста пластид с развитой гранулярно-ламеллярной структурой начинается образование ферментов, участвующих в процессе фотосинтеза. Так, один из основных специфических ферментов, катализирующий первую реакцию тем-новой фазы фотосинтеза,— Rubisco присутствует в проростках еще до того, как листья приобретают зеленую окраску. Однако на свету происходит новообразование этого фермента. Для начала фотосинтеза необходимо также наличие акцептора углекислого газа рибулезо-1,5-бифосфата (РБФ). Это соединение образуется как промежуточный продукт апотомического пути дыхательного распада. Еще до прорастания в зародыше присутствует ключевой фермент апотомического пути дыхания — глюкозофосфатдегидрогеназа. В процессе прорастания активность этого фермента резко возрастает, что приводит к первичному накоплению акцептора углекислоты — РБФ. Затем начинает работать цикл Кальвина и активность глюкозофосфатдегидрогеназы падает.
Как было видно, прорастание семян зависит от условий. Существует много механизмов, регулирующих этот процесс, и множество барьеров, препятствующих ему, причем у разных видов эти механизмы и эти барьеры специфичны. Такая сложная система выработалась в процессе эволюции, и дает определенные преимущества растительному организму. В любой естественной обстановке все условия, необходимые для прорастания всех видов растения и даже всех семян, данного вида не могут быть соблюдены. Это приводит к тому, что прорастание семян даже одного и того же вида растягивается на несколько лет. Последнее дает возможность сохранить определенное количество жизнеспособных семян в том случае, если неблагоприятные условия привели к гибели проросшей особи. Именно поэтому прорастание семян является критическим этапом жизни растительного организма, во многом обеспечивающим выживаемость того или иного вида.
Дата добавления: 2015-06-05; просмотров: 2273;