ДРУГИЕ ФАКТОРЫ, ОГРАНИЧИВАЮЩИЕ ДИАПАЗОН ДВИЖЕНИЯ
Ряд факторов могут ограничивать диапазон движения сустава. Это может быть мышечный дисбаланс, неадекватный мышечный контроль, возраст мышцы или ее иммобилизация.
Мышечный дисбаланс. Здоровые мышцы сохраняют структурный гомеостаз. Ключом такого структурного баланса является взаимная тяга мышц-антагонистов, расположенных на противоположной стороне сустава (рис. 3.13). Дисбаланс этих сил тяги может повлиять на диапазон движения. Мышечный дисбаланс может быть обусловлен рядом факторов, включая наличие гипертонических мышц (т. е. мышц, находящихся в состоянии контрактуры или спазма) или слабых мышц.
Неадекватный мышечный контроль. Даже если человек от рождения наделен достаточной гибкостью, локальный мышечный контроль может быть неадекватным для выполнения определенных элементов, требующих проявления гибкости. Это объясняется тем, что многие из них состоят из дополнительных компонентов. В данном контексте мы рассматриваем мышечный контроль как наличие адекватного баланса, координации или контроля за частью (частями) тела или достаточной силы для выполнения конкретного элемента, требующего проявления гибкости. Например, чтобы выполнить такой элемент, как «ласточка», требуется умение сохранять равновесие. Кроме того, необходимо обладать достаточной силой, чтобы принять и сохранить необходимое положение. Выполнение более сложных двигательных умений требует соответствующего сочетания всех необходимых качеств и компонентов, подготовленности.
Влияние процесса старения на мышцы. Естественный процесс старения ведет к снижению функциональных возможностей мышцы. Если человек ведет малоподвижный образ жизни, страдает какими-нибудь заболеваниями и т. д., этот процесс протекает намного быстрее. С физиологической
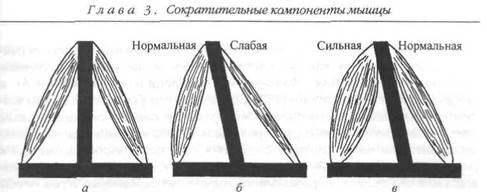
Рис. 3.13. Мышечный баланс: а — мышечный баланс обеспечивает структурный баланс;
б— дисбаланс нередко наблюдается при ослаблении одной мышцы; в — иногда дисбаланс
обусловлен тем, что одна из мышц оказывается слишком сильной (Wakher, 1981)
точки зрения наиболее очевидным дегенеративным изменением, связанным с процессом старения, является постепенная атрофия мышечной массы. Она обусловлена снижением размера и количества мышечных волокон (Grob, 1983; E.Gutmann, 1977; Hooper, 1981). Как замечает Уилмор (1991), «... это снижение количества саркомеров может обусловливать снижение подвижности, обычно наблюдаемое у пожилых людей». Отмечаются значительные колебания возраста, в котором проявляются эти изменения, существенно варьируется и степень изменений. С возрастом также сокращается количество нервных клеток в скелетно-мышечной системе (E.Gutmann, 1977).
По мере атрофии мышечных волокон происходит их замена жировой и волокнистой (коллагеновой) тканью. Коллаген — главный компонент соединительной ткани — характеризуется исключительно низкой эластичностью. Поэтому вполне очевидно, что даже незначительное увеличение количества коллагена в мышце приводит к существенному снижению ее подвижности. Это явление изучали Альнакиб, Аль-Заид и Голд-спинк (1984), использовав для этого камбаловидную мышцу и длинный разгибатель пальцев крыс. Их исследования подтвердили, что общее содержание коллагена с возрастом постоянно увеличивается. Кроме того, они обнаружили в молодой мышце более низкую интенсивность развития пассивного напряжения на каждую единицу увеличения длины. В мышце взрослого животного пассивное напряжение развивалось с большей интенсивностью.
«Данные, касающиеся тугоподвижности мышц и соединительной ткани, характеризуются корреляцией, за исключением данных о камбаловидной мышце старых животных. Пассивное механическое поведение мышцы непосредственно связано с концентрацией коллагена».
Таким образом, эти изменения в определенной степени обусловливают возрастное снижение гибкости.
Влияние иммобилизации. Способность мышцы адаптировать свою длину в зависимости от положения продемонстрировал в экспериментах
Наука о гибкости
 еще Марви (Marvey, 1887). В последние 20 лет механизмы адаптации длины мышцы изучали как на клеточном, так: и на ультраструктурном уровнях. Исследования, проведенные Голдспинком (1968, 1976) и П.Уильямсом и Голдспинком (1971), показали, что увеличение длины мышечного волокна во время нормального развития связано со значительным увеличением количества саркомеров вдоль длины волокон. Так как длина филаментов актина и миозина постоянна, адаптация взрослой мышцы к различной функциональной длине, по-видимому, должна включать производство или удаление определенного количества саркомеров с тем, чтобы сохранить правильную длину саркомера относительно всей мышцы (Goldspink, 1976; ГаЬагу и др., 1972).
еще Марви (Marvey, 1887). В последние 20 лет механизмы адаптации длины мышцы изучали как на клеточном, так: и на ультраструктурном уровнях. Исследования, проведенные Голдспинком (1968, 1976) и П.Уильямсом и Голдспинком (1971), показали, что увеличение длины мышечного волокна во время нормального развития связано со значительным увеличением количества саркомеров вдоль длины волокон. Так как длина филаментов актина и миозина постоянна, адаптация взрослой мышцы к различной функциональной длине, по-видимому, должна включать производство или удаление определенного количества саркомеров с тем, чтобы сохранить правильную длину саркомера относительно всей мышцы (Goldspink, 1976; ГаЬагу и др., 1972).
При иммобилизации камбаловидной мышцы взрослой кошки в удлиненном положении мышца адаптируется к новой длине. Табари с коллегами (1972) обнаружили, что это удлинение осуществляется за счет появления около 20 % новых саркомеров. П.Е.Уильямс и Голдспинк {! 973) обнаружили, что новые саркомеры добавляются на концах имеющихся мио-фибрилл. В случае денервации и иммобилизации в удлиненном состоянии производится около 25 % саркомеров (Goldspink и др., 1974). После прекращения иммобилизации «нормальная» и денервированная мышца быстро восстанавливают свою первоначальную длину (Goldspink и др., 1974; Tabary и др., 1972). Совсем недавно было проведено исследование с целью определить, способна ли внеглазная мышечная система трех обезьян адаптироваться таким образом, как мышцы конечности кошек (A.B.Scott, 1994). Исследования показали увеличение длины глазных мышц на 18, 25 и 33 % вследствие наложения швов.
При иммобилизации конечности с мышцей в сокращенном положении мышечные волокна теряют до 40 % саркомеров (Tabary и др., 1972). При денервации и иммобилизации й сокращенном положении наблюдали снижение числа саркомеров на 35 % (Goldspink и др., 1974). Однако затем количество саркомеров восстанавливается по мере восстановления длины мышцы (Gofdspink и др., 1974; Tabary и др., 1972).
Таким образом, адаптация количества саркомеров в зависимости от функциональной длины мышц происходит не под нейронным контролем. Скорее всего, реакция на количество пассивного напряжения, которому подвергается мышца, носит миогенный характер (Goldspink, 1976; P.E.Wiliiams, Goldspink, 1976).
Наряду со снижением длины волокон, а также количества и длины саркомеров ученые выявили и уменьшение растяжимости (увеличение пассивного сопротивления) мышц, иммобилизованных в сокращенном положении (Goldspink, 1976; Goldspink, Williams, 1979). Этот процесс происходил независимо от того, была ли мышца денервирована или нет (Goldspink, I976). Голдспинк и Уильяме (1979) также обнаружили, что потеря соединительной ткани происходит с меньшей интенсивностью, чем потеря сократительной ткани мышцы. Таким образом, относительное количество соединительной ткани увеличивается (Goldspink, 1976; Goldspink, Williams, 1979). Кроме того, П.Е. Уильяме и Голдслинк (1984) установили, что волокна коллагена в иммобилизованной мышце распола-
Глава 3 Сокртитепьные компоненты мышцы
 гаются под более острым углом к оси мышечных волокон, чем в обычной мышце. Это изменение, очевидно, влияет на эластичность мышцы.
гаются под более острым углом к оси мышечных волокон, чем в обычной мышце. Это изменение, очевидно, влияет на эластичность мышцы.
Снижение степени растяжимости, вероятно, является защитным механизмом, который не допускает резкого перерастяжения мышцы (Goldspink, 1976; Goldspink, Williams, 1979). Этот механизм особенно важен для сокращенной мышцы (т. е. мышцы, утратившей саркомеры), поскольку растягивание даже в обычном диапазоне движения приведет к перерастяжению сарко-меров и филаменты миозина и актина не смогут осуществить взаимное перекрытие, что вызовет временное повреждение мышцы (Goldspink, 1976; Tabary и др., 1972). С другой стороны, изменения в эластичных свойствах мышцы, иммобилизованной в удлиненном положении, не произойдут, поскольку адаптация обратно пропорциональна, а вероятность перерастяжения мышцы такая же, как и для обычной мышцы (Tabary и др., 1972).
Однако указанное выше снижение степени растяжимости не является просто защитной функцией. Главным последствием изменений количества саркомеров и длины мышцы является смещение кривой длины-напряжения мышцы влево (при иммобилизации мышцы в сокращенном положении) или вправо (при иммобилизации в удлиненном положении). Эти изменения длины помогают мышце адаптироваться к генерированию оптимальных уровней напряжения при новом положении и новой длине.
На основании приведенных выше результатов П.Е.Уильямс с коллегами (1988) решили определить, обусловливает ли недостаточное растяжение или недостаточная сократительная активность сокращение количества саркомеров, увеличение концентрации коллагена и увеличение тугопод-вижности мышц, которое наблюдается при их иммобилизации в сокращенном положении. Они обнаружили, что аккумуляция соединительной ткани, имеющая место в иммобилизованных мышцах, может быть предотвращена либо путем пассивного растяжения, либо путем активной (электрической) стимуляции.
В связи с этим возникает вопрос, являются ли короткие периоды растягивания достаточно эффективными для предотвращения изменений в соединительной ткани мышц, длине волокон и количестве саркомеров, т. е. способствуют ли они сохранению диапазона движений сустава? Исследование, проведенное П.Е. Уильямсом (1988) на мышах, показало, что периоды пассивного растягивания продолжительностью всего 15 мин (через день) обеспечивают нормальные пропорции соединительной ткани. В то же время они не способствуют предотвращению снижения длины мышечных волокон, которое ведет к заметному ограничению диапазона движения.
Дата добавления: 2015-05-19; просмотров: 1897;