ДВИГАТЕЛЬНЫЕ СИСТЕМЫ СТВОЛА МОЗГА
В этот отдел мозга физиологи включают продолговатый мозг, мост и средний мозг.
А. Ядра ствола мозга. Здесь находятся ядра Ш-ХП пар черепных нервов: глазодвигательного нерва (п. осЫотогогшз, III); блокового нерва (п. хгосЫеапз, IV); отводящего нерва
ПО
(п. аЬаисепз, VI), все они управляют движением глаз в различных направлениях; тройничного нерва (п. гп^егшпиз, V), иннервирую-щего жевательную мускулатуру; лицевого нерва (п. [аааПз, VII), двигательное ядро которого вызывает сокращение мимической мускулатуры; преддверно-улиткового нерва (п. уезИЫососЫеаш, VIII) - вестибулярные его ядра получают импульсы от вестибулорецеп-торов, участвуют в регуляции позы и равновесия тела, улитковые ядра иннервируют слуховые рецепторы; языкоглоточного нерва (п. §1о55орЬагуп§еи5, IX), содержащего двигательные, чувствительные и секреторные парасимпатические волокна (стимулируют секрецию слюны околоушной*железы); блуждающего нерва (п. уа^из, X), регулирующего функции внутренних органов (парасимпатический нерв); двигательное ядро добавочного нерва (п. ассеззогшз, XI), вызывающее сокращение грудинно-ключично-сосцевидной и трапециевидной мышц; двигательное ядро подъязычного нерва (п. Ьуро§1о55из, XII) иннервирует мышцы языка. Б. Двигательные системы продолговатого мозга и моста. Из перечисленных ядер продолговатый мозг включает ядра 1Х-ХП пар черепных нервов, а также образования ретикулярной формации. Мост представлен ядрами V—VIII пар черепных нервов и также ретикулярной формацией. В стволе мозга проходят восходящие
и нисходящие нервные пути. Важную роль в регуляции тонуса мышц туловища и конечностей играют вестибулярные ядра, основным из которых является ядро Дейтерса.
1. Рефлексы, осуществляемые продолговатым мозгом и мостом, можно объединить в три основные группы: 1) жизненно важные вегетативные (дыхания, сердечно-сосудистой системы, пищеварительной системы); 2) защитные рефлексы - чихания, кашля, мигания, слезоотделения, рвотный; 3) соматические рефлексы, участвующие в регуляции тонуса и двигательной активности мышц туловища, конечностей, шеи, лица. Рефлексы любого отдела ЦНС изучают после отделения вышележащих отделов мозга. У бульбарного животного (сделана перерезка ствола мозга между мостом и средним мозгом) сохраняются все рефлексы, осуществляемые спинным мозгом, а также указанными черепными нервами и их ядрами. Особо важную роль в регуляции тонуса мышц тулови-ща и конечностей играет ядро Дейтерса. С помощью этого ядра осуществляется, в частности, лабиринтный рефлекс позы - повышение тонуса мышц-разгибателей при положении животного вверх ногами. Рефлекс в натуральных условиях направлен на сохранение позы. Он возникает с рецепторов вестибулярного аппарата (при его разрушении рефлекс исчезает). Проприорецепторы мышц шеи при изучении этого рефлекса должны быть выключены, что дости-" гается путем гипсования шеи (исключаются сгибание шеи и раздражение проприорецепторов мышц шеи).
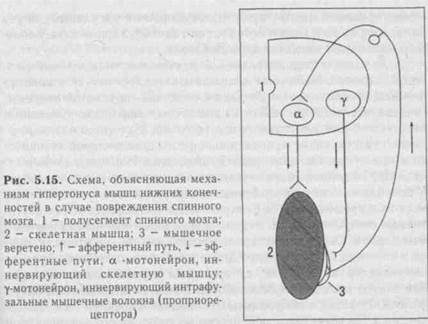
В. Двигательные системы среднего мозга включают красные ядра, передние и задние холмики четверохолмия: передние -первичные зрительные центры вместе с латеральными коленчатыми телами; задние - первичные слуховые центры, вместе с медиальными коленчатыми телами, ядра III и IV пар черепных нервов, черную субстанцию, ретикулярную формацию, голубое пятно. Рефлексы, осуществляемые средним мозгом (мезенцефаль-ное животное) более совершенны, их набор значительно больше, тонус мышц близок к нормальному, поскольку сохраняется тормозящее влияние красного ядра на ос- и у-мотонейроны спинного мозга, наблюдается незначительная ригидность мышц (рис. 5.9). Соматические рефлексы, связанные с изменением положения тела (точнее, головы) и перемещением его в пространстве Р.Магнус разделил соответственно на две группы - статические и статоки-нетические.
Статические рефлексы (перераспределение мышечного тонуса при изменении положения головы), в свою очередь, делятся на две группы: рефлексы позы (шейные и лабиринтные вестибулярные) и выпрямительные (выпрямление головы и туловища). Реф-
леке выпрямления головы (если, например, животное лежит на боку) возникает с рецепторов вестибулярного аппарата, кожи и сетчатки. При этом возбуждаются проприорецепторы мышц шеи и начинается выпрямление туловища. При выключении рецепторов кожи или вестибулярного аппарата выпрямительные рефлексы не нарушаются. При выключении обеих названных рефлексогенных зон выпрямительные рефлексы не возникают. Для осуществления выпрямительных рефлексов, т.е. восстановления нарушенной позы, необходимо обязательное участие красных ядер, их разрушение в эксперименте предотвращает возникновение выпрямительных рефлексов. При изменении положения головы возникают также рефлексы компенсаторного положения глаз, которые обеспечивают правильное видение при различных положениях головы относительно тела и в пространстве. Например, если повернуть голову вправо, глазное яблоко поворачивается влево, как бы стремясь удержать первоначальное поле зрение.
Статокинетические рефлексы возникают при ускорениях прямолинейного и вращательного движений организма. Перераспределение тонуса направлено на преодоление действующих на человека ускорений и сохранение нормальной позы. Для их осуществления необходимо обязательное участие структур среднего мозга.
Примером рефлекса прямолинейного ускорения является лифтный рефлекс. В начале быстрого подъема лифта (или остановки движущегося вниз лифта) в нижних конечностях повышается тонус мышц-разгибателей. В начале опускания лифта (или остановки движущегося вверх лифта) повышается тонус сгибателей (имеется и иная точка зрения о причине сгибания и разгибания конечностей при спуске и подъеме лифта). О рефлекторном характере изменения тонуса мышц конечностей при лифтном рефлексе свидетельствует его отсутствие у животных при выключении вестибулярных рецепторов разрушением лабиринта.
Кроме регуляции мышечного тонуса, позы и равновесия структуры ствола головного мозга участвуют в контроле спинального автоматизма шагания и, следовательно, в контроле локомоции (например, ходьбы). Произвольная локомоция без участия корковых нейронов невозможна.
Функции голубого пятна. Оно оказывает моделирующее влияние на двигательные механизмы спинного мозга. Аксоны норадре-нергических нейронов голубого пятна идут в переднем и боковом канатиках к а-мотонейронам передних рогов спинного мозга. Полагают, что норадреналин является тормозным медиатором для а-мотонейронов.
Функции черной субстанции. Она участвует в регуляции мышечного тонуса, координации жевания и глотания, мелких движений конечностей (например, печатание на машинке, письмо), в осуществлении эмоций. В случае повреждения черной субстанции наблюдается бедность движений.
Во всех двигательных реакциях и регуляции мышечного тонуса весьма важную роль играет ретикулярная формация, основная масса нейронов которой находится в стволе мозга.
Дата добавления: 2015-05-16; просмотров: 1036;