Механизмы, регулирующие кровообращение
Кровообращение в мозгу нельзя рассматривать изолированно от кровообращения всего организма в целом, так как усиленная работа тех или иных органов или тканей постоянно сопровождается перераспределением крови в организме. Интенсивная работа какого-либо органа вызывает приток крови к нему; в то же время поступление крови к другим органам уменьшается. При таких перераспределениях крови по всему организму мозг по степени кровенаполнения может находиться в синергических или антагонистических отношениях с рядом других органов.
Находясь в общей системе с другими органами, мозг в то же время занимает среди них особое положение. В отличие от остальных органов и тканей мозг всегда активен, что в свою очередь является результатом непрерывного функционирования нервных клеток. Определенный уровень процессов обмена веществ, протекающих в основных элементах нервной ткани, требует постоянного притока кислорода или, иначе говоря, непрерывного поступления крови к мозгу. В соответствии с только что сказанным существует целый ряд условий и механизмов, обеспечивающих постоянство кровоснабжения мозга. К числу их должно быть отнесено нахождение мозга в герметически закрытом окостеневшем черепе, сифоны на артериях, снабжающих головной мозг, при вхождении их в полость черепа, синокаротидный механизм и другие.
Среди механизмов, регулирующих поступление крови к мозгу, различают приспособления, ограждающие его от чрезмерного переполнения кровью и, напротив, обеспечивающие подачу большого количества крови з случае повышенной деятельности мозга. Повышение активности мозга сопровождается вступлением в действие как механизмов, ведающих перераспределением крови во всем организме, так и механизмов, перераспределяющих кровь в пределах только мозга. Деятельность последних обеспечивает работающим отделам мозгового вещества значительно большее количество крови по сравнению с участками, функциональная деятельность которых в данный момент заторможена.
Механизм действия приспособлений, принимающих участие в регуляции мозгового кровообращения, недостаточно установлен. Правильнее было бы сказать, что наши знания о нем находятся на самой начальной ступени их решения.
Прежде чем перейти к рассмотрению регуляции кровообращения в мозгу посредством экстрацеребрально расположенных механизмов, необходимо остановиться на регуляции кровообращения в организме вообще. Необходимость эта диктуется тем, что кровоснабжение мозга, являющегося частью целого организма, находится в тесной зависимости от изменений величины общего кровяного давления.
Великому русскому физиологу И. П. Павлову принадлежит заслуга установления зависимости деятельности сердечно-сосудистой системы от влияний со стороны центральной нервной системы. Если до И. П. Павлова имелись только отдельные указания о возможности участия нервной системы в регуляции кровяного давления, то после целого ряда экспериментальных исследований И. П. Павлова в 70—90-х годах прошлого столетия роль нервной системы при выполнении данной функции организма стала несомненной. Исследования И. П. Павлова, посвященные изучению регуляции сердечной деятельности и регуляции тонуса сосудистой стенки, сыграли огромную роль, так как они дали совершенно новое направление изучению кровообращения. Поэтому понятно, что в дальнейшем проблема кровообращения организма стала исследоваться под углом влияния нервной системы на деятельность всей кровеносной системы.
Удержание кровяного давления на постоянном уровне является основным для нормального организма. Экспериментальные доказательства этого положения, как известно, получены И. П. Павловым в 1879 г. В опытах И. П. Павлова было показано, что кормление собак сухим хлебом, нагрузка организма водой (кормление мясной похлебкой) не сопровождаются заметными колебаниями общего кровяного давления. Выводы из многочисленных опытов И. П. Павлова категорически опровергали существовавшие до этого предположения о возможности значительных колебаний кровяного давления в нормальных условиях. И. П. Павлову принадлежит также заслуга экспериментального изучения факторов, служащих для поддержания постоянного уровня кровяного давления в организме. Опыты с нарушением анатомической и физиологической целости блуждающих нервов, перерезкой или сильным охлаждением их позволили И. П. Павлову сделать заключение, что блуждающий нерв является одним из регуляторов общего кровяного давления (1882,1883). Результаты целого ряда исследований привели И. П. Павлова к утверждению решающей роли центральной нервной системы в регулировании и удержании постоянного уровня кровяного давления в организме. Раскрыв в своих экспериментах часть механизмов, обеспечивающих определенный уровень кровяного давления, И. П. Павлов считал задачей будущих исследователей дальнейшую разработку этой проблемы. «Огромная важность точного изучения приспособлений, стоящих на страже этого стремления к постоянству, неизмерима. Ближайшее проникновение в механику иннервации кровообращения обещает, кроме того, получение сведений о некоторых, до сих пор упущенных из виду, раздражителях, благодаря которым и пускаются в ход то та, то другая деятельность. Только достигнув этого, мы сможем говорить как о внешних, так и о внутренних влияниях, управляющих кровообращением. До сих пор все притязания практической медицины на управление кровообращением остаются одними лишь пожеланиями»1.
1 И. П. Павлов, Полное собрание трудов, изд. Академии наук СССР, т. I, стр. 54, 1940.
Представления о регуляции кровообращения складывались очень медленно. Первые шаги в этом направлении были сделаны Клод Берна-ром, Людвигом и др. В экспериментах этих исследователей раздражение спинного мозга вызывало повышение общего кровяного давления к одновременное сужение сосудов. Напротив, перерезка спинного мозга сопровождалась падением кровяного давления и расширением сосудов в органах и тканях, расположенных ниже уровня перерезки [Шифф (Schiff), 1855].
Таким образом, была установлена зависимость просвета сосудов от величины и изменений общего кровяного давления. Однако механизм этого явления оставался неясным. Можно было лишь предполагать, что области, из которых исходят влияния, повышающие тонус сосудов, расположены выше уровня спинного: мозга.
Открытию сосудодвигательного центра предшествовал ряд умозрительных предположений, указывавших на возможность существования его в продолговатом мозгу. Так, например, Клод Бернар объяснял причину повышения содержания сахара в крови при уколе в продолговатый мозг повреждением вазомоторного центра печени. Отдельные исследователи считали вероятным расположение в продолговатом мозгу общего сосудодвигательного центра, регулирующего кровообращение во всем организме (Шифф). П. Н. Овсянников (1873) представил, наконец, четкие экспериментальные доказательства существования этого центра в определенном месте продолговатого мозга. Рядом последовательных перерезок мозгового ствола он установил границы сосудодвигательного центра в мозгу кролика. Согласно данным П. Н. Овсянникова, центр, тонизирующий сосуды, расположен на 2 мм ниже четверохолмия и на 3—4 мм выше писчего пера. Этот центр представляет собой парное образование, каждая половина которого отделена от средней линии ромбовидной ямки на 2,5 мм.
Пытаясь соотнести сосудодвигательные реакции с ядерными образованиями продолговатого мозга, В. М. Бехтерев высказал мнение, что наиболее вероятным было бы считать сосудодвигательным центром нижнее центральное ядро. В дальнейшем зона локализации этого центра была расширена В. М. Бехтеревым. Он включил в нее ретикулярную субстанцию, расположенную вокруг нижнего центрального ядра. Последующие исследователи в своих экспериментальных работах не смогли указать на вазомоторную функцию ни одного из ядерных скоплений продолговатого мозга (Бруштейн, 1901; Рэнсон и Монье, 1939; Лима, 1939; Склярский, 1940; Александер, 1946, и др.). Благодаря этому обстоятельству в настоящее время большинство исследователей предполагает, что местом расположения сосудодвигательного центра является серая ретикулярная субстанция продолговатого мозга (substantia reticularis grisea).
На более точную локализацию сосудодвигательного центра в продолговатом мозгу указывает, к сожалению, пока только одна работа. Бэкер (Baker, 1950) с сотрудниками представил результаты исследований ретикулярной субстанции продолговатого мозга людей, умерших от бульбарного полиомиэлита. В тех случаях, когда у больного отмечались нарушения со стороны сердечно-сосудистой системы, поврежденными оказывались значительные по размерам клетки, расположенные в средне-нижнем отделе ретикулярной субстанции.
Клинические симптомы высокого или, напротив, низкого кровяного давления, неправильный ритм сердечной деятельности, увеличение частоты пульса и плохое наполнение его и т. д. появлялись при разрушении структуры более чем 19% нервных клеток в указанной области. При
дальнейшем развитии процесса нарушения кровообращения прогрессировали. Давление падало до низкого уровня, пульс не прощупывался, кожа становилась холодной и липкой, затем следовал бред, кома и гипертермия и, наконец, наступала смерть.
На основании патологоанатомического изучения мозга людей, погибших при явлениях расстройства сердечно-сосудистой деятельности, Бэ-кер заключил, что сосудодвигательный центр локализуется в средне-нижней части ретикулярной субстанции продолговатого мозга.
В состав сосудодвигательного центра входят нервные клетки моторного типа с длинными, многократно ветвящимися дендритами. Размер этих клеток весьма различен. Так, длина клеток колеблется от 75 до 35 м, а ширина — от 35 до 20 м. Клетки описанного «центра» разнообразны по форме и могут быть продольно вытянутыми, сферическими, веретенообразными или угловатыми. Ядра клеток пузырькообразны, ядрышко в ядре расположено эксцентрически. Тигроид вариирует по форме от тонкогранулярного до комкообразного, особенно по периферии клетки, и нитевидного. Располагаясь в ретикулярной субстанции между медиальной петлей и моторным ядром блуждающего нерва (п. ambiguus), клетки сосудодвигательного центра впервые появляются на уровне верхнего полюса подъязычного нерва и могут быть прослежены вплоть до верхнего полюса ядра лицевого нерва, Иначе говоря, клеточная группа распределяется на протяжении приблизительно 14 мм.
Полученные Бэкером данные требуют не только дополнительных патологоанатомических исследований на подобном же материале, но и специальной экспериментальной проверки. Подтверждение этих данных даст возможность говорить о локализации сосудодвигательного центра в продолговатом мозгу в цитоархитектоническом отношении уже с полной уверенностью.
Трудность получения точной цитоархитектоничсской локализации сосудодвигательного центра в эксперименте объясняется, как нам кажется, особой организацией этих центров. Обычно понятие центра совпадает с понятием ядра, т. е. под центром понимают обособленную группу клеток, близко прилежащих друг к другу, выполняющих одну какую-нибудь функцию. Примером этому могут служить ядра черепномозговых нервов, красное ядро, ядра гипоталамуса и др.
Как уже было сказано, территория, занятая ядром, обычно характеризуется скоплением большого количества капилляров, резко выделяющих данное ядро среди окружающих образований (рис. 84, а, б, в, г). В самом ядре капилляры оплетают каждую нервную клетку, а иногда даже внедряются в ее протоплазму.
При сравнительном изучении капиллярной сети и клеточного строения ретикулярной субстанции продолговатого мозга приходится признать существование несколько другой организации центра, не совпадающей с понятием клеточного ядра.
Если обратиться к строению ретикулярной субстанции, в которой на основании физиологических и клинических данных предполагается локализация дыхательного и сосудодвигательного центра, то обнаружить в ней особые клеточные группы невозможно.
Здесь имеются разрозненно лежащие гигантские клетки моторного типа, отличающиеся от обычных моторных клеток наличием многочисленных очень длинных дендритов, переходящих через среднюю линию даже на противоположную сторону. Около каждой из этих клеток располагается несколько более мелких нервных клеток с короткими ветвящимися дендритами. Капиллярная сеть в ретикулярной субстанции очень
однообразна и состоит из более или менее одинаковых по величине петель. Следовательно, центры продолговатого мозга имеют структуру, резко отличающую их от центров других отделов мозга.
На основании результатов экспериментов целого ряда авторов было установлено, что раздражение некоторых участков центральной нервной системы выше уровня продолговатого мозга в свою очередь сопровождается изменениями общего кровяного давления (В. Я. Данилевский, 1874; В. М. Бехтерев, 1905; Гесс, 1938, и др.). Эти высшие вегетативные центры, регулирующие кровообращение ib отдельных участках организма, располагаются в гипоталамической области промежуточного мозга.
В 1874 г. В. Я. Данилевский впервые показал существование влияний на сердечную деятельность и сосудистую систему со стороны полушарий головного мозга. При раздражении коры в области коркового представительства лицевого нерва, а также при раздражении частей задней крестовидной извилины В. Я. Данилевский отметил изменение деятельности сердца и значительные колебания кровяного давления. Величина изменений находилась в прямом отношении к продолжительности и интенсивности раздражения. Повышение кровяного давления в некоторых опытах от 150 до 240 мм и изменение ритма работы сердца продолжались 6—12 секунд. Результаты работ В. Я. Данилевского впоследствии были не только подтверждены, но и расширены замечательным русским ученым В. М. Бехтеревым и его учениками. Так, еще в 1881—1885 гг. В. М. Бехтерев в совместной работе с Н. Миславским установил, что раздражение внутренней части передней и задней крестовидных извилин и наружной поверхности височной доли мозга собак сопровождается резким повышением кровяного давления. Напротив, понижение кровяного давления, в дальнейшем, правда, сменяющееся повышением, может быть получено при раздражении током наружной и средней части крестовидной извилины. Депрессорный эффект наблюдается и при раздражении теменной области полушария.
Таким образом, сосудодвигательный эффект оказалось возможным получить с довольно обширных областей мозговой коры. Неэффективными в этом отношении оказались лишь передние отделы лобной доли и затылочная доля. Изменение ритма сердечной деятельности наблюдалось в опытах В. М. Бехтерева и Н. Миславского при раздражении передней области мозговой коры и белого вещества передней крестовидной извилины. Обширные экспериментальные исследования и многочисленные клинические наблюдения привели В. М. Бехтерева к выводу о существовании в коре полушарий головного мозга сердечных и сосудодви-гательных центров, осуществляющих свое влияние на соответствующие, расположенные ниже, центры посредством специальных проводников.
Поскольку в эксперименте было возможно вызвать сосудодвигательный эффект для отдельных частей организма, В. М. Бехтерев считал возможным говорить о наличии в коре сосудодвигательных центров руки, ноги, лица и т. д.
Данные В. М. Бехтерева и Н. Миславского вскоре были воспроизведены в вышедшей из лаборатории В. Я. Данилевского в 1892 г. работе
A. Черевкова, а затем в работах Минора, Тарханова и др. Изучение
влияния на сердечно-сосудистую деятельность коры головного мозга в
процессе ее развития, произведенное Готье (1903) в лаборатории
B. М Бехтерева, показало, что изменение сосудистого давления и ритма
работы сердца при раздражении коры могут быть отмечены у щенков
начиная только со второго месяца внеутробной жизни.
Влияние коры на сердечно-сосудистую деятельность при раздраже-
нии различных участков ее, установленное в эксперименте, в дальнейшем было подтверждено учениками В. М. Бехтерева (Сорохтин, Л. Л. Васильев и Подерни) методом сочетательных рефлексов. Аналогичные результаты тем же методом условных рефлексов получил Цитович (1918) в лаборатории И. П. Павлова.
В настоящее время вопрос о регулирующем влиянии коры головного мозга на сердечно-сосудистую деятельность подвергся всестороннему изучению в лаборатории К. М. Быкова.
Начиная с 1927 г. и до настоящего времени сотрудники К. М. Быкова проводят детальное изучение изменения просвета сосудов конечностей под влиянием условнорефлекторной деятельности коры. Сочетая деятельность условных раздражителей (света, звонка, ударов метронома) с безусловным тепловым, Холодовым или химическим раздражением в целом ряде работ, А. А. Рогов (1928, 1932, 1933, 1941), А. Т. Пшонник (1939, 1947, 1948), К. М. Быков и А. Т. Пшонник (1949) установили значительное влияние коры на сосуды.
Самые разнообразные раздражители (звук, прикосновение и т. д.) дают ответные реакции со стороны сосудов. Вместе с тем условнореф-лекторная реакция сосудов теснейшим образом связана с состоянием нервной системы, так как утомительная физическая или умственная работа ведет к резкому понижению или исчезновению условных рефлексов со стороны сосудов.
Не менее тщательные исследования, предпринятые в лаборатории К. М. Быкова в отношении изучения влияния коры головного мозга на сердце, позволили установить, что все проявления деятельности сердца находятся под непосредственным воздействием коры. Иначе говоря, миокард и проводящая система сердца испытывают постоянное регулирующее воздействие с коры (В. Е. Делов, 1939; Е. Г. Петрова, 1942; Г. А. Самарин, 1942; Левитин, 1944; К. М. Смирнов, 1941, и т. д.). Открытое А. И. Смирновым корковое представительство блуждающего нерва дает возможность говорить о влиянии коры на все стороны сердечнососудистой системы.
Изучение влияния со стороны нервной системы на сосуды показало, что кровеносные сосуды снабжены волокнами, по которым к ним поступают сосудосуживающие и сосудорасширяющие импульсы. Сосудосужи-ватели, или вазоконстрикторы, являются нервами симпатической природы. Преганглионарные волокна сосудосуживателей берут начало от клеток боковых рогов спинного мозга в грудном и поясничном его отделе, идут в составе передних корешков и оканчиваются на клетках ганглиев пограничного столба.
От клеток ганглиев начинаются постганглионарные волокна, оканчивающиеся в мышечных элементах сосудов. Электрофизиологические данные указывают, что сосудосуживающие нервы являются путями проведения импульсов, поддерживающих сосудистый тонус. При отведении токов действия с данных нервов получают постоянные осцилляции, исчезающие после перерезки вазоконстрикторов одновременно с расширением сосудов области, иннервируемой тем или иным нервом.
В отношении происхождения сосудорасширяющих нервов окончательно установленных данных не существует. Часть из сосудорасширяющих волокон имеет несомненную парасимпатическую природу. Речь идет о нервных волокнах, идущих к сосудам в составе черепномозговых, а также крестцовых нервов. В последнее время получены данные, согласно которым часть сосудорасширителей принадлежит к симпатическому отде-
лу вегетативной червной системы. Сосудорасширяющие нервные волокна, имеющие указанный источник происхождения, идут к сосудам в составе симпатического нерва. Наконец, нужно упомянуть о существовании точки зрения, указывающей на возможность проведения сосудорасширяющих импульсов по обычным афферентным нервам задних корешков спинного мозга по типу аксон-рефлексов. Если сосудорасширители парасимпатической и симпатической природы являются проводниками влияний, исходящих из центральной нервной системы, то сосудорасширители, если они действительно идут ib составе задних корешков, должны служить для осуществления сосудистых реакций, имеющих местное значение.
Помимо двигательных волокон, сосуды снабжены также чувствительными волокнами. Последние являются путями проведения барохемо-рецептивной, а также болевой чувствительности от сосудов в центральную нервную систему. Помимо этих, диффузно рассеянных по всем сосудам рецепторов, большие скопления их сосредоточиваются на внутренней стенке ряда крупных сосудов. Такого рода своеобразные чувствительные зоны располагаются в начальной части дуги аорты, в полых венах (в участках их, прилежащих к предсердию), а также в так называемом каротидном синусе.
Основным физиологическим раздражителем рецепторов в указанных зонах является растяжение сосудистой стенки под влиянием увеличения кровяного давления. Возникающие в этих случаях импульсы идут по афферентным нервам особого порядка в продолговатый и промежуточный мозг к расположенным там сердечно-сосудистым центрам. В результате изменения состояния этих центров меняется сердечная деятельность и тонус сосудистой стенки и происходит повышение или понижение общего кровяного давления.
Так, например, усиленный приток венозной крови к сердцу влечет за собой растяжение полых вен и раздражение рецепторов, заложенных в их устье. Импульсы, возникающие в указанном рецептивном поле, возбуждают центр симпатических нервов сердца - в шейной части спинного мозга и угнетают деятельность центра блуждающего нерва в продолговатом мозгу. В результате этого увеличивается число сердечных сокращений и минутный объем крови. При растяжении аорты увеличивающимся давлением крови раздражаются заложенные в стенке ее рецепторы. Афферентный импульс поступает к сердечно-сосудистым центрам продолговатого мозга и вызывает рефлекторное понижение кровяного давления, возвращающегося к нормальному уровню.
Открытию синокаротидной рецепторной зоны предшествовала работа многих исследователей. Было собрано много разрозненных фактов, прежде чем были установлены характер и путь рефлексов, возникающих при раздражении расположенных в ней нервных окончаний. Так, Чер-мак в 1866 г. получил замедление пульса, падение кровяного давления и углубление дыхания при надавливании на боковую поверхность шеи в области верхней трети ее. Он объяснил этот эффект раздражением блуждающего нерва в результате сдавления сосудисто-нервного пучка. Однако дальнейшие исследования не подтвердили этого предположения. Вскоре после наблюдения Чермака было отмечено, что замедление сердечной деятельности возникает только при давлении на область бифуркации сонной артерии.
В последующем Пагано (Pagano, 1900) показал, что замедление пульса может быть вызвано лишь при увеличении давления в общей сонной артерии. Исходя из этого, он пришел к выводу о существовании какого-то нервного механизма, расположенного в месте деления сонной
|
артерии на внутреннюю и наружную ветви. По мысли Пагано, этот механизм раздражался при увеличении давления в сонной артерии, в результате чего осуществлялось рефлекторное замедление сердечной деятельности и мозг предохранялся от слишком сильного повышения давления. Особая роль сонных артерий в регуляции кровообращения во всем организме была установлена также в опытах Сичилиано (Siciliano, 1900). Сичилиано подтвердил, что нарушение дыхательной деятельности в довольно резкой форме может наблюдаться при перевязке обеих общих сонных артерий ниже места их разветвления, тогда как перевязка обеих позвоночных артерий никогда не влечет за собой изменения дыхания.
Характерные явления, сопровождающие зажатие или перевязку сонных артерий, послужили поводом для целого ряда физиологических и морфологических работ, которые ставили своей целью выяснить физиологическое значение и морфологическое строение области разветвления сонной артерии.
Так, например, Геринг (1923) удалял участок сонной артерии в области ее деления на внутреннюю и наружную ветви или перерезал нервную ветвь, идущую от места бифуркации в языкоглоточный нерв. В обоих случаях наблюдалось повышение кровяного давления. В другой серии опытов при раздражении центрального конца перерезанной ветви языкогло-точного нерва было получено падение кровяного давления и замедление сердечной деятельности.
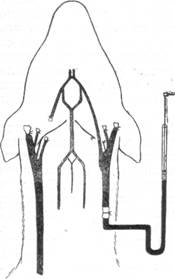
| Рис. 104. Схема опыта Моисеева. |
Таким образом, экспериментами Геринга был установлен путь чувствительного импульса от рецепторов внутренней
стенки бифуркации к продолговатому мозгу. Ветвь языкоглоточного нерва, служащая для проведения этих импульсов, получила название синусного нерва. Область бифуркации сонной артерии, являющаяся местом сосредоточения нервных окончаний указанного нерва, вошла в литературу под названием каротидного синуса.
Дальнейшее систематическое изучение функций каротидных синусов стало возможным благодаря методу, предложенному Е. А. Моисеевым (1927), работавшим в лаборатории Н. Н. Аничкова. Разработанный им метод заключался в том, что вслед за перевязкой всех периферических ветвей сонной артерии создавалась своеобразная культя, связанная с центральной нервной системой синокаротидным нервом (рис. 104). Меняя давление в культе, Е. А. Моисеев имел возможность наблюдать чистый эффект от той или другой манипуляции. При такой постановке опыта повышение давления в культе постоянно имело своим следствием падение кровяного давления, иногда сопровождавшееся даже прекращением сердечной деятельности.
Описанный метод исследования настолько полно соответствовал целям изучения реакций каротидного синуса, что все последующие работы в основном были проведены при некоторых видоизменениях или дополнениях по методу Моисеева.
В результате многочисленных исследований было установлено, что основной функцией синокаротидной зоны является понижение общего кровяного давления в ответ на повышение давления в синусе [Гейманс (Heymans), 1933, и многие другие]. Реакция возникает вследствие раздражения сложно построенных рецепторных окончаний синусного нерва, заложенных в стенке каротидного синуса [де Кастро (de Castro), 1926; Н. Г. Смирнова, 1948, и т. д.]. Афферентной дугой при осуществлении рефлекторного понижения давления является, как уже говорилось, синусный нерв, проводящий импульсы к центрам сосудисто-сердечной деятельности в продолговатом мозгу. Под влиянием этих импульсов возникает торможение активности сосудодвигагельного центра и повышение активности центра блуждающего нерва. Эфферентная часть рефлекторной дуги к сердцу представлена блуждающим нервом. Эфферентные импульсы направляются также в спинной мозг к находящимся там сосудосуживающим центрам и оттуда по симпатическим сосудосуживающим нервам к сосудам внутренностей, конечностей и кожи. Результатом торможения является падение общего кровяного давления и замедление дыхательной деятельности. При длительном повышении давления в ка-ротидном синусе может наступить даже остановка дыхания.
При уменьшении давления в общей сонной артерии, что в свою очередь сопровождается снижением давления на рецепторы синокаротидной зоны, реакция протекает по-иному. Сосудодвигательный центр в этих случаях высвобождается от постоянных влияний со стороны синокаротидной зоны. Повышение активности сосудодвигательного центра влечет за собой значительно большее, чем обычно, возбуждение симпатических сосудистых центров спинного мозга. Повышение тонуса этих центров ведет к сужению сосудов брюшной полости, конечностей и кожи. Одновременно происходит усиление сердечной деятельности вследствие возбуждения симпатического центра сердца. Все это ведет к повышению общего кровяного давления, а также учащению и увеличению дыхательных движений, а иногда и к одышке (гиперпноэ).
Таким образом, рефлексы с каротидного синуса вызывают не только понижение общего кровяного давления, но могут способствовать и повышению давления. Поэтому синокаротидная зона играет чрезвычайно важную роль в регуляции кровообращения во всем организме.
Как показали многочисленные исследования, помимо барорецепто-ров, регистрирующих колебания давления, в синокаротидной зоне имеются и хеморецепторы, реагирующие на изменения химического состава крови [де Кастро, 1926; А. А. Смирнов и др.]. Хеморецепторы каротидного синуса располагаются около бифуркации и сосредоточены в особом образовании, носящем название glomus caroticus. Так же как и баро-рецепторы, нервные окончания отмечающие изменение химизма крови, не являются специфическими только для синокаротидной зоны. Такого рода рецепторные окончания описаны и для дуги аорты, где они располагаются в paraganglion aorticum.
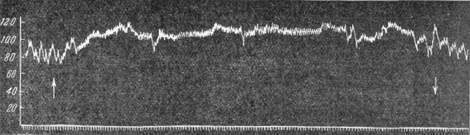
Функция описываемого рода рецепторов может быть обнаружена при введении в ток крови различных веществ (Гейманс и др., 1931; П. Н. Веселкин, 1937; С. Асратян, А. А. Кузнецов, 1938, и др.). Введение гипотонического раствора поваренной соли или дестиллированной воды в общую сонную артерию (рис. 105) сопровождается повышением общего кровяного давления. Такая же реакция со стороны кровяного давления наблюдается при увеличении содержания углекислоты в крови или сдвиге реакции в кислую сторону. Напротив, увеличение щелочности крови имеет своим следствием падение кровяного давления. Па-
дение кровяного давления вызывает также введение вток крови физиологического раствора (рис. 106). Фармакологические вещества также действуют на хеморецепторы различно, вызывая то возбуждение, то торможение их. Афферентные пути хеморецепторных окончаний идут независимо от афферентных проводников барорецепторов. Так, после перерезки нерва, идущего от каротидного синуса, исчезают реакции в ответ на изменения давления в синусе, но сохраняются реакции на изменения химического состава крови. Эти реакции исчезают только в условиях полной денервации каротидного синуса.
|
| Рис. 105. Повышение кровяного давления при введении в общую сонную артерию дистиллированной воды. |
Таким образом, при изменении величины общего кровяного давления или химического состава крови прессорные и депрессорные реакции
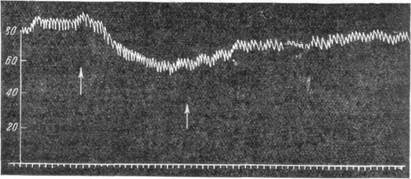
Рис. 106. Понижение кровяного давления при введении в обшую сонную артерию физиологического раствора.
являются несомненно функцией синокаротидной зоны. Вместе с тем экспериментальные данные указывают, что в условиях нормальной жизнедеятельности организма, когда существуют колебания давления только при систоле и диастоле, основной функцией синокаротидной зоны является торможение активности сосудосуживающего центра.
Постоянная циркуляция крови через каротидный синус создает непрерывное состояние возбуждения в барорецепторах, расположенных в стенке сонной артерии в месте бифуркации ее, и возбуждение хеморе-цепторов glomus caroticus.
Это обстоятельство с полной очевидностью выступает при электрофизиологической регистрации токов действия с неповрежденного синусного нерва [Бронк (Bronk), 1931, и др.].
При отведении токов действия с указанного нерва в физиологических условиях отмечается непрерывное проведение афферентных импульсов. от синокаротидной зоны к продолговатому мозгу. Как видно из рис. 107, при каждой систоле наблюдается групповой разряд импульсов, тогда как при диастоле по нерву идут лишь нерегулярные импульсы разнообразной амплитуды. Изучение афферентной импульсации одного концевого аппарата каротидного синуса кролика показало, что при бы-стром повышении давления во время систолы рецептор дает около 55 импульсов в секунду [Бронк и Стелла (Bronk a. Stella), 1932]. Падение давления влечет за собой уменьшение числа импульсов в единицу времени. При низком и среднем давлении разряд иногда прекращается во зремя диастолы. Так, например, при колебаниях давления от 40 до 15 мм.
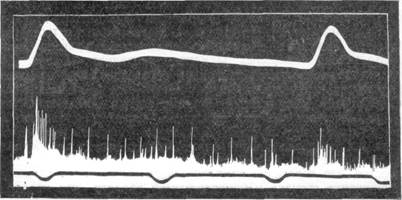
Рис. 107. Токи действия в неповрежденном синусном нерве в условиях нормальной жизнедеятельности организма.
Верхняя запись — кровяное давление; нижняя запись — токи действия в синусном нерве. Видно увеличение ритма и амплитуды импульсов во время систолического подъема давления.
ртутного столба концевой аппарат или совсем не дает разрядов, или дает несколько осцилляции во время систолы. Повышение давления сопровождается усилением импульсации. Во время систолы разряды становятся продолжительнее, ритм делается более частым, амплитуда отдельных колебаний увеличивается. При достижении кровяным давлением высокого уровня разряды могут сделаться непрерывными, слабо варии-рующими по частоте.
Сопоставление данных электрофизиологического анализа с результатами, полученными при повышении или понижении давления в каротидном синусе, позволяет отметить, что увеличение силы и продолжительности разрядов концевых аппаратов влечет за собой понижение кровяного давления. Напротив, ослабление импульсации имеет своим следствием прессорную реакцию повышения общего кровяного давления. Точно так же электрическое раздражение синусного нерва сопровождается понижением общего кровяного давления, а перерезка или кокаини-зация — повышением кровяного давления.
Иначе говоря, всякое усиление рефлекторных влияний со стороны синокаротидной зоны на сосудосуживающий центр в продолговатом мозгу ведет к понижению общего кровяного давления, т. е. к торможению функциональной деятельности этого центра. В то же время ослабление рефлекторных влияний при выключении баро- и хеморецепторов—понижением давления в каротидном синусе или при введении в него дестил-
лированной воды находит свое отражение в повышении давления крови в организме. Повышение давления в этом случае указывает на прояв-.ление деятельности сосудосуживающего центра, освобожденного от тормозящих влияний со стороны синокаротидной зоны.
Аналогичные данные получены и при изучении влияний, испытываемых сосудосуживающим центром со стороны рецепторной зоны, расположенной в области дуги аорты. Отведение токов действия от депрессор-ного нерва показало, что при повышении давления в данной области имеет место переход залповой формы импульсации в непрерывную. При этом одновременно происходит понижение кровяного давления вследствие расширения сосудов и ослабления работы сердца (П. К. Анохин и Я. И. Шумилина, 1947).
Таким образом, экспериментальные данные указывают, что синока-ротидная и кардиоаортальные рецепторные зоны представляют собой источник тормозящих влияний, умеряющий функциональную деятельность сосудодвигательного центра. Но в таком случае естественно уместен вопрос о факторах, создающих определенный уровень процессов жизнедеятельности в нервных элементах указанного центра. Другими словами, возникает вопрос о причинах, обеспечивающих настолько высокую активность сосудодвигательного центра, что для понижения ее организм выработал специальные приспособления. Выяснение условий, создающих постоянно высокую активность сосудодвигательного центра, является до настоящего времени чрезвычайно сложной задачей в связи с чем при многообразии мнений определенного ответа на поставленный выше вопрос пока дать не представляется возможным.
Известно, что перерезка симпатического нерва имеет своим следствием расширение сосудов в иннервируемой области. Это указывает на существование определенного влияния со стороны симпатической части вегетативной нервной системы на гладкую мускулатуру сосудистой стенки. Следовательно, можно считать, что периферический тонус сосудистой стенки создается импульсами, поступающими по симпатическим нервам. Вместе с тем установлено, что перерезка спинного мозга на различных уровнях под продолговатым ведет к резкому падению кровяного давления до 60—20 мм ртутного столба. В связи с результатами описанных выше опытов возникло предположение, что сосудистый тонус и высота кровяного давления всецело поддерживаются и регулируются симпатической нервной системой. Основной центр последней располагается в продолговатом мозгу и своей деятельностью обеспечивает жизненно необходимый уровень кровяного давления, равный 80—120 мм ртутного столба.
Но в последующих экспериментах с использованием более совершенных методических приемов было обнаружено, что при поддержании искусственного дыхания перерезка спинного мозга под продолговатым не влечет за собой резкого падения давления. Это обстоятельство указывало на ограничение роли сосудодвигательного центра в создании и поддержании сосудистого тонуса во всем организме. Отсюда возникло заключение, что определенный уровень кровяного давления не зависит в полной мере от влияний со стороны этого центра, но обеспечивается и какими-то механизмами другого порядка.
Кроме того, удаление у кошек симпатических узлов в шейной, груд-ной и пояснично-сакральной области показало, что и периферический сосудистый тонус до известной степени независим от центров, располагающихся в спинном мозгу. Но если разобщение высших сосудодвига-тельных центров в продолговатом и спинном мозгу от сосудистойсистемы
202
не вело к резким изменениям уровня кровяного давления, то регуляция его была отчетливо нарушена. Животные с удаленными симпатическими цепочками были мало жизнеспособны и могли существовать лишь в хороших лабораторных условиях [Кеннон (Cannon), 1929; Кеннон и Брайт (Cannon a. Bright), 1931].
И. Н. Канторович и Г. П. Конради (1948) вводили новокаин в ток кро-ви животного и отмечали лишь незначительное падение кровяного давления. Поскольку новокаин снимает действие симпатической системы, эти исследователи сделали заключение о существовании на периферии какого-то механизма, поддерживающего тоническое состояние сосудистой стенки. Возникающий под влиянием этих механизмов сосудистый тонус, по их мнению, служит фоном, на котором проявляется действие симпатической нервной системы как регулятора тонического состояния гладкой мускулатуры. Канторович и Конради не отрицают существования сосудодви-гательного центра в продолговатом мозгу, они лишь отвергают за ним функцию поддержания тонуса гладкой мускулатуры сосудистой стенки.
Однако согласиться с подобной точкой зрения на роль сосудодвига-тельного центра нельзя. Хотя перерезка спинного мозга под продолговатым не во всех случаях сопровождается падением давления до критического уровня, это не противоречит еще концепции о тонизирующем влиянии сосудодвигательного центра. Перерезка спинного мозга на уровне VII шейного сегмента или применение в том же участке новокаина постоянно приводит к падению кровяного давления на 35—40 мм ртутного столба. Следовательно, кровяное давление при перерезке спинного мозга до того или иного уровня всегда падает, что не может не указывать на прекращение влияний со стороны сосудодвигательного центра. Уровень давления после перерезки будет различным в зависимости от высоты его перед началом опыта.
Сохранение кровяного давления на некотором, необходимом для поддержания жизни животного, уровне говорит о том, что тоническое состояние гладкой мускулатуры сосудов создается не только импульса-ми, исходящими от сосудодвигательного центра в продолговатом мозгу. В поддержании этого тонуса принимают участие нижележащие симпатические центры, расположенные на различных уровнях спинного мозга, симпатические узлы, а также и сама сосудистая стенка. Но, несмотря на то, что периферический сосудистый тонус поддерживается совместным действием целого ряда механизмов, среди них роль сосудодвигательного центра очень значительна. Вопрос о факторах, обеспечивающих тоническое состояние самого сосудодвигательного центра, не утратил своего значения.
По аналогии с механизмами тонуса поперечнополосатой мускулатуры приходится думать о существовании каких-то «сторонних» рецепторов, влияющих на постоянную активность сосудодвигательного центра. Как известно, тонус поперечнополосатой мускулатуры поддерживается не только проприоцепторами, располагающимися в самих мышцах или по соседству с ними в сухожилиях мышц и суставах, но и «сторонними» рецепторными приборами, такими, как, например, вестибулярный аппарат. Но как при адэкватном, так и неадэкватном раздражении последнего наблюдается также падение кровяного давления (Б. К. Клосов-ский и Н. В. Семенов, 1947; Шпигель и др.). В наших опытах совместно с Н. В. Семеновым было показано, что при исключении влияния со стороны полушарий головного мозга всякое раздражение вестибулярного аппарата имеет своим следствием падение давления на 20—30 мм ртут-ного столба, продолжающееся 30 секунд (рис. 108, а, б). Отсюда можно
сделать вывод, что «стороннее» действие вестибулярного аппарата выражается в тормозящем влиянии на сосудодвигательный центр. В литературе до настоящего времени не описано ни одного достоверного факта, который говорил бы о наличии нервного, активно тонизирующего влия ния какого-либо рецепторного аппарата на сосудодвигательный центр.
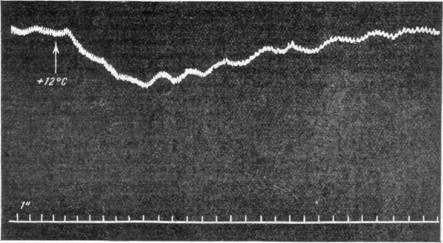
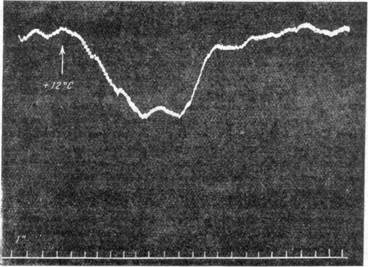
Рис. 108. Падение кровяного давления при раздражении вестибулярного аппарата.
Дата добавления: 2015-05-16; просмотров: 1107;