Спинальные рефлексы
Важную роль в развитии представлений о рефлекторной деятельности спинного мозга сыграли открытия и обобщения английского физиолога, лауреата Нобелевской премии Чарлза Шеррингтона (1859-1952).
Объем функций, осуществляемых спинным мозгом, чрезвычайно велик. В нем находятся центры всех двигательных рефлексов (за исключением мускулатуры головы), всех рефлексов мочеполовой системы и прямой кишки, рефлексов, обеспечивающих терморегуляцию, регулирующих метаболизм тканей, центры большинства сосудистых рефлексов, центр сокращения диафрагмы и др. В естественных условиях эти рефлексы всегда испытывают влияние высших отделов головного мозга.
Степень проявления рефлексов зависит от того, сохраняются ли связи структур спинного мозга со структурами головного мозга. После децеребрации (удаления головного мозга) или спинализации (отделения спинного мозга от головного посредством перерезки) исчезают многие сложные формы активности, создаваемые спинным мозгом. При этом определенное значение принадлежит уровню организации подопытного животного. Например, спинальная лягушка, представитель низших позвоночных, может сидеть и вырываться, когда ее схватывают. Спинальная же собака сама не может ни стоять, ни ходить. Это объясняется тем, что разобщение спинного мозга и вышележащих структур нарушает рефлекторные дуги, ответственные за осуществление определенных реакций. При этом, в частности, исчезают периодические разряды дыхательных мышц, обеспечивающие дыхательные движения, пропадают тонические разряды симпатических нейронов, поддерживающих сосудистый тонус и соответственно артериальное давление.
Как правило, рефлексы спинальных животных являются координированными. Возбуждение у них каждой группы рецепторов сопровождается своим специфическим ответом. Например, механическое раздражение кожи ступни у лягушки вызывает сгибание раздражаемой конечности и разгибание другой. Раздражение рецепторов мочевого пузыря и прямой кишки сопровождается рефлекторным сокращением их мускулатуры.
В связи с отсутствием тонических влияний структур головного мозга изменяется функциональное состояние и самих спинно-мозговых нейронных систем. К числу таких нарушений относится прекращение сложных локомоторных актов типа шагания. Характерно, что исчезновение этих актов после спинализации удается восстановить введением животному веществ, способствующих выделению медиаторов синаптическими окончаниями перерезанных путей.
В зависимости от числа нейронов, участвующих в проведении возбуждения, рефлекторные дуги спинного мозга делятся на моносинал-тические и полисинаптические. Моносинаптическая дуга состоит из чувствительного нейрона с рецепторами мышечных веретен и эффекторного нейрона, оканчивающегося на мышечных волокнах. Классическим примером моносинаптической дуги может служить рефлекторная дуга коленного рефлекса, в которой возбуждение от рецептора до эффектора проходит всего за 0,5-1,0 мс, т. е. время, необходимое для прохождения возбуждения всего лишь через один синапс.
В полисинаптической дуге на пути возбуждения от рецептора к эффектору помимо чувствительного и эффекторного нейрона находятся еще вставочные нейроны. Таким образом, возбуждение в этой дуге проходит не через один, а через несколько синапсов, определяющих время латентного периода ответа и суммарной синаптической задержки. При осуществлении рефлекторных реакций в моно- и полисинаптических дугах вовлекаются также координирующие рефлекс многочисленные интернейроны.
Подобный анализ периферических воздействий, а также надсст-ментарный контроль за рефлекторной деятельностью спинного мозга происходит с помощью длинных многоэтажных рефлекторных дуг. Их центры локализованы в подкорковых областях и коре головного мозга.
К числу рефлексов спинного мозга относятся защитные рефлексы, рефлексы на растяжение, мышц-антагонистов, висцеромоторные, вегетативные рефлексы. Эта классификация весьма условна, и весь ее смысл в том, что она указывает на многообразие рефлекторных ответов. Даже у спинального животного трудно встретить рефлексы, которые относились бы только к одной из названных групп.
Раздражение в виде укола кожи стопы вызывает у лягушки защитный рефлекс – либо отдергивание лапки при легком уколе, либо вовлечение в реакцию другой лапки и отодвигание от источника при более сильном воздействии, наконец, убегание животного при значительном болевом раздражении, когда в возбуждение вовлекаются многие структуры нервной системы.
Рефлексы растяжения проявляются укорочением мышцы в ответ на ее растяжение. Основными рецепторами в этом случае служат нервно-мышечные веретена, афферентным звеном – чувствительные волокна соматических нервов и дорсальных корешков спинного мозга. Эти рефлекторные дуги чаще всего замыкаются в спинном мозге. Начало и конец рефлекторной дуги связаны с мышцей. Рефлексы наиболее выражены в мышцах-разгибателях. Для того чтобы организм мог противостоять силе земного притяжения, эти мышцы должны находиться в состоянии тонического напряжения. Биологическое значение этих рефлексов состоит в том, что они участвуют в сохранении статики и положения тела, регулируя степень сокращения мышцы в соответствии с падающими на нее раздражениями. Особое значение этот вид рефлексов имеет у копытных животных, хотя и у человека они хорошо развиты.
Рефлексы мышц-антагонистов лежат в основе локомоторных актов и характеризуются тем, что при возбуждении мотонейронов сгибателей одновременно происходит торможение мотонейронов мыщц-разгибателей. При этом в конечности другой стороны наблюдаются обратные явления. В целом это создает правильное чередование противоположных по функциональному значению мышечных сокращений. Механизм, обусловливающий такое чередование активности различных двигательных ядер, например при ходьбе, локализуется в интернейронном аппарате спинного мозга. Вместе с тем для его активации необходимо поступление тонического нисходящего сигнала из двигательных центров головного мозга.
Висцеромоторные рефлексы возникают при возбуждении афферентных волокон внутренних органов и характеризуются появлением двигательных реакций мышц грудной и брюшной стенки, мышц-разгибателей спины. Возникновение этих рефлексов связано с существованием конвергенции висцеральных и соматических афферентных волокон к одним и тем же интернейронам спинного мозга.
Вегетативные рефлексы заключаются, во-первых, в появлении полисинаптических разрядов в преганглионарных симпатических волокнах в ответ на возбуждение симпатических и соматических чувствительных клеток, во-вторых, в возникновении рефлекторных реакций парасимпатических нейронов в ответ на раздражение чувствительных путей. Наряду с выполнением собственных рефлекторных реакций нейронные структуры спинного мозга служат аппаратом для реализации большого числа сложных процессов, осуществляемых различными отделами головного мозга. Это управление может быть прямым, когда нисходящие пути непосредственно связаны с мотонейронами спинного мозга, и непрямым через интернейроны, образующие короткие межсегментарные связи. Свойства последних и особенности связей с нисходящими волокнами и моторными нейронами создают возможность к интеграции поступающих сигналов, их переработке и пространственному перераспределению.
Проводниковые функции спинного мозга
Помимо рефлекторной деятельности еще одной важной функцией спинного мозга является проведение импульсов. Оно осуществляется белым веществом, состоящим из нервных волокон.
В результате эволюционного развития простая рефлекторная дуга, лежащая в основе функции нервной системы, усложняется и в каждой ее части вместо одного нейрона возникают цепи нервных клеток, аксоны которых образуют проводящие пути. Под проводящими путями принято понимать группы нервных волокон, характеризующиеся общностью строения и функций. Они связывают различные отделы спинного мозга или спинной и головной мозг. Все нервные волокна одного пути начинаются от однородных нейронов и заканчиваются на нейронах, выполняющих одинаковую функцию.
В соответствии с функциональными особенностями различают ассоциативные, комиссуральные и проекционные (афферентные и эфферентные) нервные волокна. Ассоциативные волокна или их пучки осуществляют односторонние связи между отдельными частями спинного мозга. Связывая разные сегменты, они образуют собственные пучки, являющиеся частью сегментарного аппарата спинного мозга. Комиссуральные волокна соединяют функционально однородные противоположные участки разных отделов спинного мозга. Проекционные волокна связывают спинной мозг с вышележащими отделами. Эти волокна образуют основные проводящие пути, которые представлены восходящими (центростремительными, афферентными, чувствительными) и нисходящими (центробежными, эфферентными, двигательными) путями.
Восходящие проводящие пути. Эти пути несут импульсы от рецепторов, воспринимающих информацию из внешнего мира и внутренней среды организма. В зависимости от вида чувствительности, которую они проводят, их делят на пути экстеро-, проприо-и интероцептивной чувствительности. Нисходящие пути передают импульсы от структур головного мозга к двигательным ядрам, осуществляющим ответные реакции на внешние и внутренние раздражения.
Основными восходящими путями спинного мозга являются тонкий пучок, клиновидный пучок, латеральный и вентральный спиноталамические тракты, дорсальный и вентральный спинно-мозжечковые тракты (рис.18.).
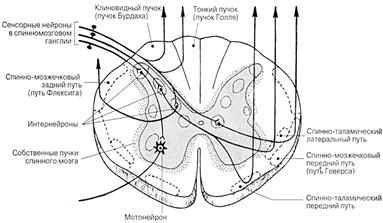
Рис. 18. Расположение восходящих проводящих путей
спинного мозга
Тонкий пучок (Голля) и клиновидный пучок (Бурдаха) составляют задние столбы спинного мозга. Эти пучки волокон являются отростками чувствительных клеток спинальных ганглиев, проводящих возбуждение от проприоцепторов мышц сухожилий, частично тактильных рецепторов кожи, висцерорецепторов. Волокна тонкого и клиновидного пучков миелинизированные, они проводят возбуждение со скоростью 60-100 м/с. Короткие аксоны обоих пучков устанавливают синаптические связи с мотонейронами и интернейронами своего сегмента, длинные же направляются в продолговатый мозг. По пути они отдают большое число ветвей к нейронам вышележащих сегментов спинного мозга, образуя, таким образом, межсегментарные связи.
По волокнам тонкого пучка проводится возбуждение от каудальной части тела и тазовых конечностей, по волокнам клиновидного пучка – от краниальной части тела и грудных конечностей. В спинном мозгу оба этих пути идут, не прерываясь и не перекрещиваясь, и оканчиваются в продолговатом мозгу у одноименных ядер (Голля и Бурдаха), где образуют синаптическое переключение на второй нейрон. Отростки второго нейрона направляются к специфическим ядрам таламуса противоположной стороны, образуя тем самым, своеобразный перекрест. Здесь они переключаются уже на третий нейрон, аксоны которого достигают нейронов IV слоя коры больших полушарий.
Считают, что по этой системе проводится информация тонко дифференцированной чувствительности, позволяющая определить локализацию, контур периферического раздражения, а также его изменения во времени.
По латеральному спиноталамическому тракту проводится болевая и температурная чувствительность, по вентральному спиноталамическому – тактильная. Существуют сведения, что по этим путям возможна также передача возбуждения от проприо- и висцерорецепторов. Скорость проведения возбуждения в волокнах составляет 1-30 м/с. Спиноталамические тракты прерываются и перекрещиваются либо на уровне сегмента, в который они только что вступили, либо вначале проходят несколько сегментов по своей стороне, а затем переходят на противоположную. Отсюда идут волокна, оканчивающиеся в зрительных буграх. Там они образуют синапсы на нервных клетках, аксоны которых направляются в кору больших полушарий.
Полагают, что по системе волокон этих путей в основном передается информация о качественной природе раздражителей.
Дорсальный спинно-мозжечковый тракт, или пучок Флексига – филогенетически это наиболее древний чувствительный путь спинного мозга. Местом расположения нервных клеток, аксоны которых образуют волокна этого пути, является основание дорсального рога спинного мозга. Не перекрещиваясь, путь достигает мозжечка, где каждое волокно занимает определенную область. Скорость проведения по волокнам спинно-мозжечкового пути около 110 м/с. По ним проводится информация от рецепторов мышц и связок конечностей. Наибольшего развития этот путь достигает у копытных животных.
Вентральный спинно-мозжечковый тракт, или пучок Говерса, также образуется аксонами интернейронов противоположной стороны спинного мозга. Через продолговатый мозг и ножки мозжечка волокна направляются к коре мозжечка, где занимают обширные площади. Импульсы со скоростью проведения до 120 м/с идут от сухожильных, кожных и висцерорецепторов. Они участвуют в поддержании тонуса мышц для выполнения движений и сохранения позы.
Нисходящие пути связывают высшие отделы ЦНС с эффекторными нейронами спинного мозга. Основными из них являются пирамидный, красноядерно-спинномозговой, вестибулоспинальный и ретикулоспинальный тракты.
Пирамидный тракт образован аксонами клеток двигательной зоны коры больших полушарий. Направляясь к продолговатому мозгу, эти аксоны отдают большое число коллатералей структурам промежуточного, среднего, продолговатого мозга и ретикулярной формации. В нижней части продолговатого мозга большая часть волокон пирамидного пути переходит на противоположную сторону (перекрест пирамид), образуя латеральный пирамидный тракт. В спинном мозгу он располагается в боковом столбе. Другая часть волокон идет, не перекрещиваясь, до спинного мозга и только на уровне сегмента, в котором оканчивается, переходит на противоположную сторону. Это прямой пирамидный тракт. Оба они заканчиваются на мотонейронах передних рогов серого вещества спинного мозга. Состав волокон этого пути неоднороден, в нем представлены миелинизированные и немиелинизированные волокна разного диаметра со скоростями проведения возбуждения от 1 до 100 м/с.
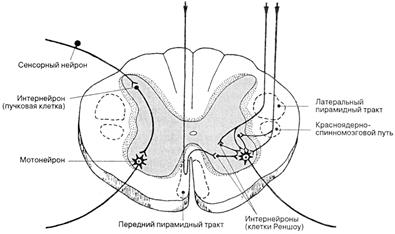
Рис. 19. Расположение нисходящих проводящих путей
спинного мозга
Основной функцией пирамидных путей является передача импульсов для выполнения произвольных движений. Надежность в осуществлении этой функции повышается благодаря дублированию связи головного мозга со спинным посредством двух путей - перекрещенного и прямого. В эволюционном ряду пирамидный тракт развивался параллельно с развитием коры больших полушарий и достиг наибольшего совершенства у человека.
Руброспинальный тракт (Монакова) образован аксонами клеток красного ядра среднего мозга. Выйдя из ядра, волокна полностью переходят на противоположную сторону. Часть из них направляется в мозжечок и ретикулярную формацию, другие – в спинной мозг. В спинном мозгу волокна располагаются в боковых столбах перед перекрещенным пирамидным путем и оканчиваются на интернейронах соответствующих сегментов. Руброспинальный тракт несет импульсы от мозжечка, ядра вестибулярного нерва, полосатого тела.
Основное назначение руброспинального тракта - управление тонусом мышц и непроизвольной координацией движений. В процессе эволюции этот путь возник рано. Большое значение он имеет у животных, слабее развит у человека.
Вестибулоспинальный тракт образован волокнами, которые являются отростками клеток ядра Дейтерса, лежащего в продолговатом мозгу. Этот тракт имеет наиболее древнее эволюционное происхождение. По нему передаются импульсы от вестибулярного аппарата и мозжечка к мотонейронам вентральных рогов спинного мозга, регулирующие тонус мускулатуры, согласованность движений, равновесие. При нарушении целостности этого пути наблюдаются расстройства координации движений и ориентации в пространстве.
В спинном мозгу помимо основных длинных имеются и короткие нисходящие пути, соединяющие между собой его отдельные сегменты.
УчебныЙ вопрос № 3
Дата добавления: 2015-05-13; просмотров: 2040;