Механизм шагательного рефлекса
Способ вызова рефлекса.Рефлекс вызывается однократным раздражением кожи конечности. Он выражается в сгибании этой конечности с одновременным дополнительным разгибанием противоположной задней конечности – перекрестный разгибательный рефлекс. Затем согнутая конечность разгибается, опускается вниз, разогнутая – сгибается и поднимается вверх и т.д. Конечности опоры не касаются (собака зафиксирована в станке с помощью лямок) и без дополнительного раздражения продолжают ритмично поочередно сгибаться и разгибаться, т.е. «шагать» в воздухе. Рефлекс может продолжаться часами, так как устранено влияние коры большого мозга. При неповрежденных сегментах, иннервирующих передние конечности (С5–Th2), и более длительном раздражении задней конечности в рефлекс вовлекаются также и передние конечности. Это наблюдение показывает наличие межсегментарной связи на уровне спинного мозга, играющей важную роль в ритмических движениях конечностей. Поочередное сокращение и расслабление мышц-сгибателей и разгибателей осуществляется в результате взаимодействия процессов возбуждения и торможения в соответствующих центрах спинного мозга под влиянием импульсов, поступающих в мозг от проприорецепторов.
Роль проприорецепторов в осуществлении шагательного рефлексаопределяется их расположением.
Мышечные веретена (мышечные рецепторы) расположены параллельно скелетной мышце: своими концами крепятся к соединительно-тканной оболочке пучка экстрафузальных мышечных волокон при помощи напоминающих сухожилия полосок соединительной ткани длиной 0,5-1 мм. Поэтому при расслаблении (удлинении) мышцы растягиваются и мышечные рецепторы, что и ведет к их возбуждению.
Мышечный рецептор состоит из нескольких поперечнополосатых интрафузальных мышечных волокон (от лат. Fusus – веретено), окруженных соединительно-тканной капсулой. Длина интрафузальных мышечных волокон равна 4-7 мм, толщина – 15-30 мкм. Длина экстрафузальных мышечных волокон – от нескольких миллиметров до многих сантиметров, толщина – 50-100 мкм. Вокруг средней части мышечного веретена несколько раз обвивается окончание одного афферентного волокна. Многие мышечные веретена иннервируются и другим (более тонким) афферентным волокном, которое также обвивает в виде спирали мышечный рецептор, но периферические его участки. Импульсы от мышечных рецепторов возбуждают нейроны своего центра и тормозят нейроны центра-антагониста. Ближе к концам мышечного веретена подходят двигательные нервные окончания, являющиеся аксонами гамма-мотонейронов спинного мозга. Их импульсация вызывает сокращение интрафузальных мышечных волокон (мышечных волокон рецептора).
Сухожильные рецепторы (рецепторы Гольджи) заключены в соединительно-тканную капсулу и локализуются в сухожилиях скелетных мышц вблизи от сухожильно-мышечного соединения. Рецепторы представляют собой безмиелиновые окончания толстого миелинового афферентного волокна (подойдя к капсуле рецепторов Гольджи, это волокно теряет миелиновую оболочку и делится на несколько окончаний). Сухожильные рецепторы крепятся относительно скелетной мышцы последовательно, что обеспечивает их раздражение при натяжении сухожилия. Поэтому сухожильные рецепторы посылают информацию в мозг о том, что мышца сокращена (напряжено и сухожилие), а мышечные рецепторы – о том, что мышца расслаблена и удлинена. Импульсы от сухожильных рецепторов тормозят нейроны своего центра и возбуждают нейроны центра-антагониста (у мышц-сгибателей это возбуждение выражено слабее, чем у разгибателей).
Элемент шагательного рефлекса – поочередное сокращение и расслабление скелетной мышцы под влиянием импульсов от про-приорецепторов, поступающих в свой центр.
Когда мышца (сгибатель или разгибатель) расслаблена и удлинена, возбуждаются мышечные веретена, импульсы от них поступают к своим α- мотонейронам спинного мозга и возбуждают их (рис. 20, а). Далее α- мотонейроны посылают импульсы к этой же скелетной мышце, что ведет к ее сокращению. Как только мышца сократилась (рис. 20, б), возбуждение мышечных веретен прекращается или сильно ослабляется (они уже не растянуты), начинают возбуждаться сухожильные рецепторы. Импульсы от них поступают в первую очередь в свой центр в спинном мозге, но к тормозным клеткам Реншоу. Возбуждение тормозных клеток вызывает торможение α-мотонейронов этой же скелетной мышцы, вследствие чего она расслабляется. Однако ее расслабление (удлинение) ведет снова к возбуждению мышечных веретен и α-мотонейронов – мышца вновь сокращается. Вследствие ее сокращения снова возбуждаются сухожильные рецепторы и тормозные клетки в спинном мозге, что вновь ведет к расслаблению скелетной мышцы и т.д.
Мышца поочередно сокращается и расслабляется в результате поступления к ее мотонейронам импульсов от собственных рецепторов. Описанные процессы в равной степени относятся и к мышце-сгибателю, и к мышце-разгибателю. При этом расслабление скелетной мышцы запускает механизмы ее сокращения, а сокращение скелетной мышцы активирует механизмы, расслабляющие мышцу.
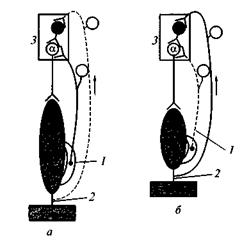
|
Рис. 20. Схема, отражающая процессы возбуждения α-мотонейронов спинного мозга и их торможения с помощью вставочных тормозных нейронов во время
сокращения и расслабления скелетной мышцы:
1 — мышечные рецепторы (мышечные веретена); 2 — сухожилия мышцы и рецепторы (рецепторы Гольджи); 3 — полусегмент спинного мозга;
а — мышца растянута и расслаблена — возбуждаются мышечные рецепторы (1);
б — мышца сокращена, напряжена и укорочена — возбуждаются сухожильные рецепторы (2); сплошные линии — импульсация выражена;
пунктирные линии — импульсация отсутствует.
Для обеспечения поочередного сгибания и разгибания конечностей при шагательном рефлексе мышцы-сгибатели и мышцы-разгибатели должны сокращаться и расслабляться последовательно друг за другом, что достигается с помощью торможения центра-антагониста при возбуждении центра-агониста. Причем если на одной ноге сокращены мышцы-сгибатели, на другой ноге должны сокращаться мышцы-разгибатели, что обеспечивается поступлением афферентных импульсов от мышечных и сухожильных рецепторов и поочередным возбуждением и торможением центров сгибателей и разгибателей. На одноименной же стороне при возбуждении центра мышцы-сгибателя тормозится центр мышцы-разгибателя.
Шагательные координированные движения у спинального животного возможны в отсутствие обратной афферентации от проприорецепторов. Они осуществляются с помощью межсегментарных связей на уровне спинного мозга. О наличии межсегментарных связей свидетельствует также факт вовлечения в шагательный рефлекс всех четырех конечностей спинальной собаки при достаточно длительном и сильном раздражении одной конечности.
Дата добавления: 2015-05-13; просмотров: 2295;