Структурні віруса
білки ®. . . . . . . . . . . ® . . . . . . . . . . . . ® . . . . . . . . . . . .
Одночасно на одній РНК матриці функціонує декілька репликаз і утворена структура називається реплікативним попередником. Приманкою для реплікації у пікорна- і калицивірусів є білок генома VРg, ковалентно пов'язаний з РНК(-) і РНК(+).
Незначна маса вірусної нуклеїнової кислоти (106-107D) призводить до того, що місткості вірусної нуклеїнової кислоти недостатньо для запису інформації про синтез усіх необхідних при циклі вірусної реплікації білків. З цією метою віруси використовують ферментний апарат клітини-хазяїна, а у ряді випадків удаються до допомоги і інших вірусів.
Наприклад, РНК вірусу-сателіта некрозу тютюну складається тільки з 1200 нуклеотидів, а кодована цією РНК білкова субодиниця включає 400 амінокислотних залишків. Очевидно, що ні для якої іншої інформації в геномі цього вірусу не вистачає місця. Тому вірус-сателліт некрозу тютюну здатний розмножуватися тільки в тих клітинах, які одночасно заражені вірусом некрозу тютюну.
Цікавим прикладом вірусів сателітів людини служить вірус гепатиту D, який репликується в гепатоцитах тільки у присутності вірусу сироваткового гепатиту В і використовує як капсид зовнішню білкову оболонку вірусу гепатиту В. Спільне розмноження вірусів гепатиту двох типів обумовлює особливо важку інфекцію з високою вірогідністю переходу в цироз печінки і в гепатокарциному.
Трансляція. Термін трансляція означає переведення генетичної інформації з матричної (інформаційної) РНК в послідовність амінокислотних залишків в білковому поліпептидному ланцюгу. Молекула іРНК через “шапочку" (кеп), розташовану на 5¢-кінці іРНК, зв'язується з малою рибосомальною субодиницею, яка рухається вздовж молекули РНК. По досягненні першого ініціюючого кодону АУГ з нею зв'язується велика рибосомальна субодиниця з транспортної РНК, і потім починається синтез білків. Трансляція триває доти, поки на рибосомі не з'явиться термінуючий кодон. Вірусні білки синтезуються на рибосомах, пов'язаних з клітинними мембранами; глікозування білків здійснюється клітинними ферментами і відбувається одночасно з синтезом поліпептиду. Синтезовані вірусні білки мігрують в різні ділянки клітини, наприклад в ядро, у випадку інфекції, обумовленої ядерними вірусами. Багато які вірусні білки модифікуються (глікозуються, ацетилюються, фосфорилюються, протеолітично нарізаються і т.д.) як в процесі трансляції, так і після неї.
Формування зрілих вірусних часток і вихід їх з клітини. Внаслідок синтезу вірусних білків і нуклеїнових кислот в клітині накопичується достатня їх кількість, і створюються умови для формування зрілих вірусних часток.
Найбільш простий механізм дозрівання віріонів у вірусів із спіральною симетрією, і вони утримують тільки один тип білків і нуклеїнову кислоту. Класичним представником таких вірусів вважається вірус тютюнової мозаїки (ВТМ). У ВТМ формування зрілої вірусної частки здійснюється шляхом самозбирання, тобто спонтанного об'єднання молекул в більш великі агрегати, в основі якого лежить специфічна взаємодія молекул білків і нуклеїнових кислот.
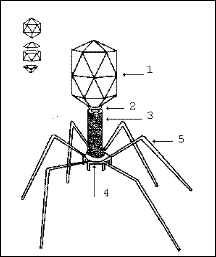
У зв'язку з різноманіттям структури вірусів різноманітними є і способи формування віріонів. У складних вірусів спочатку формуються нуклеокапсид і серцевина, які потім взаємодіють з білками вірусних оболонок. Процес формування зрілих вірусних часток у складно влаштованих вірусів можна розглянути на прикладі Т-парних фагів кишкової палички. Морфогенез Т-парних фагів здійснюється через ряд послідовних стадій. При цьому три основних компоненти фагу - головка, відросток і фібрили (рис. 23) утворюються незалежно один від одного. У складних вірусів кожен подальший етап збирання віріону ініціюється завершенням попереднього етапу.
У складних вірусів людини і тварин спочатку формується нуклеокапсид і серцевина, які потім взаємодіють з білками вірусних оболонок. Більшість оболонкових вірусів здобуває свої оболонки в ході відбруньковування через плазмолему або мембрану цитоплазматичної вакуолі. Таким чином, поліпротеїдна оболонка більшості вірусів є фрагментом плазмолеми клітини-господаря і за складом ліпідів дуже близька до неї. Виключенням є віруси віспи і гепатиту В, які невідбруньковуються, а здобувають свої зовнішні поліпротеідні оболонки іншим способом.
|
Рис. 23. Схематичне зображення віріону фага
Т4
1 - головка з ДНК-геномом; 2 - комірець; 3 - нескорочений чохол відростка, що складається з 24 витків довгої білкової молекули; 4 - базальна пластинка зі шпичками, які взаємодіють з оболонкою бактеріальної клітини; 5 - фібрили (нитки відростка фагу).
1. Утворюється головка фагу, що складається більш ніж з 10 типів білків і заповнюється довгою ниткою ДНК. Можливо, в цьому процесі беруть участь поліаміни (путресцин і спермідин), що входять до складу головки зрілого вірусу.
2. Затравкою для формування фагового відростка слугує базальна пластинка, побудована з 15 типів білків.
3. Після завершення збирання головки і відростка вони об'єднуються.
4. Нитки відростка, що складаються з продуктів 4 генів, збираються незалежно від інших вірусних структур, але прикріплюються вони до базальної пластинки тільки після прикріплення головки і відростка.
Лімітуючим фактором в збиранні вірусної частки є матриксний білок - гідрофобний білок, здатний до взаємодії як з білками вірусних оболонок, так і з ліпідами клітинної мембрани. Відразу ж після свого синтезу матриксний білок вбудовується в плазмолему з внутрішнього, цитоплазматичного боку ліпідного шару. Включення матриксного білка до плазмолеми є сигналом до збирання вірусної частки.
Синтез матриксного білка має значні відмінності в клітинах різного походження і завдяки його лімітуючій ролі в збиранні вірусної частки, він є критичним фактором, що визначає тип інфекції.
Незначна продукція матриксного білка призводить до формування абортивних інфекцій, що характеризуються відсутністю або незначною кількістю інфекційного потомства, і персистуючих інфекцій, при яких вірусспецифічні компоненти накопичуються в клітині і переходять до дочірніх клітин. Матриксний білок виявлено у вірусів грипу, сказу, ретровірусів.
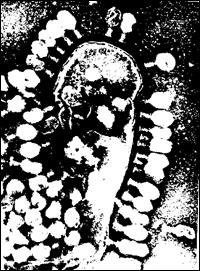
Вихід вірусів з клітини здійснюється шляхом відбруньковування і шляхом “вибуху”, при якому прості і безоболонкові віруси тварин і бактеріофаги виходять у позаклітинний простір після руйнування клітин. У руйнуванні бактерій під впливом фагів (рис. 23) бере участь фермент лізоцим, що розщеплює бактеріальні пептидоглікани.
Віруси рослин і деякі віруси людини і тварин (віруси герпесу, респіраторно-синцитіальні віруси) можуть проникати в незаражену клітину з інфікованої через плазмодесми.
Починаючи від моменту адсорбції на клітинній мембрані цикл розмноження вірусів займає від 20 - 40 хвилин (віруси бактерій) до 40-60 годин (віруси людини і тварин). Така велика різниця у часі, необхідному для вірусної репродукції, можливо пов'язана з відмінністю мікроорганізмів і клітин еукаріот і з різницею у відстані, яку повинні долати до моменту самозбирання складові елементи вірусів.
Якщо вірус передається від одного покоління клітин до іншого в ході клітинного поділу, латентний вірусний геном називається провірусом або, у разі вірусів бактерій, профагом, а клітини, які несуть такий профаг - лізогенними клітинами. Присутність геному вірусу в лізогенній культурі можна виявити при спонтанному вивільненні вірусу з клітинної популяції. У лізогенних клітинах не транскрибується жоден з вірусних генів, необхідних для вірусної інфекції. У експериментальних лізогенних культурах виявляється дуже невелика кількість вірусної іРНК, більша частина котрої синтезується на опероні, до складу якого входить ген, кодуючий синтез білків, що репресують транскрипцію решти вірусного геному. Інактивація репресора за допомогою опромінення ультрафіолетовими променями або обробки різними хімічними речовинами індукує розмноження вірусу.
|
Рис. 24. Частково лізована клітина Escherichia coli В, до поверхні якої приєднані віріони фагу Т4.
Явище лізогенії є одним з механізмів формування латентних вірусних інфекцій і лежить в основі перетворення нормальної клітки в пухлинну. Інтеграція вірусного геному в геном клітини і його подальше виключення зі складу клітинного геному створює умови для захоплення і перенесення (явище трансдукції) в нову клітину частини генетичного апарату клітину-господаря з інформацією, що не має безпосереднього відношення до життєвого циклу вірусу (явище конверсії). У ході конверсії, наприклад, бактерії роду Salmonella здобувають нові полісахаридні антигени в складі клітинної оболонки, а здатність збудників дифтерії (бактерій Corinebacterium diphtheriae) продукувати токсин залежить від бактеріофагу, що знаходиться в бактеріальній клітині в стані профагу.
Дата добавления: 2015-05-03; просмотров: 1169;