Адсорбція.
У основі адсорбції лежать два механізми:
1) неспецифічний, оборотний - віріони утримуються на поверхні клітини за допомогою електростатичних і ван-дер-ваальсових сил;
2) специфічний, в основі якого лежить взаємодія специфічних рецепторів вірусу з відповідними рецепторами клітини.
Клітинні рецептори різні для різних груп вірусів. Рецепторними функціями для ряду вірусів володіють глікопротеїди клітинних плазматичних мембран і суттєвим компонентом в них є вуглеводи. Рецептором для вірусів грипу є гли-копротеїд, що містить нейрамінову кислоту. Рецепторні функції мають також клітинні ліпіди. Клітина може бути заражена вірусом і у тому випадку, коли у неї відсутні специфічні вірусні рецептори (шляхом обробки клітини інфекційними вірусними нуклеїновими кислотами, наприклад, РНК вірусу поліомієліту; зараження вірусами клітинних протопластів; використанні "гібридних" вірусних часток, що містять нуклеїнову кислоту одного вірусу, а білкову оболонку (тобто рецептори) іншого.
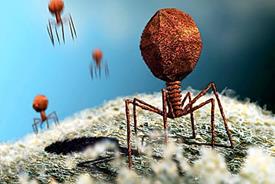
Рецептори служать не лише для прикріплення вірусів до клітинної поверхні, але і для їх подальшого транспортування усередину клітини. Вірусні рецептори можуть бути унікальними органелами, які можна побачити в електронному мікроскопі (хвостові структури у Т-фагів (рис. 11), грибоподібні вирости оболонки вірусу імунодефіциту людини) або морфологічно менш виражені структури, що складаються з глікопротеїдів на поверхні вірусних оболонок (гемаглютинін у ортоміксовірусів). Рецепторні ділянки зазвичай розташовуються на дні поглиблень і щілин на поверхні віріона, що таким чином захищає їх від блокуючої дії специфічних противірусних антитіл, діаметр яких більше діаметру щілини.
Рис. 18. Адсорбція бактеріофага Т4 на клітинній стінці кишкової палички
 На ефективність адсорбції вірусу на клітинній мембрані впливає концентрація вірусів, температура і стан клітини, наявність в середовищі електролітів (катіонів) і вільних амінокислот - кофакторів адсорбції, тканинна специфічність (рис. 18, 19).
На ефективність адсорбції вірусу на клітинній мембрані впливає концентрація вірусів, температура і стан клітини, наявність в середовищі електролітів (катіонів) і вільних амінокислот - кофакторів адсорбції, тканинна специфічність (рис. 18, 19).
Рис. 19. Адсорбція вірусів поліоми на мембрані чутливої клітини. Електронна мікроскопія. Комп'ютерна реконструкція.
 Зазвичай тваринна клітина містить близько 500000 рецепторів і на клітині можуть сорбуватись безліч віріонів. Проте, як правило, клітина стійка до повторного зараження вірусом того ж типу.
Зазвичай тваринна клітина містить близько 500000 рецепторів і на клітині можуть сорбуватись безліч віріонів. Проте, як правило, клітина стійка до повторного зараження вірусом того ж типу.
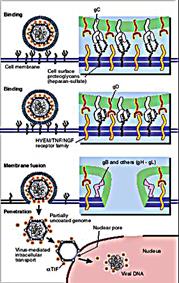
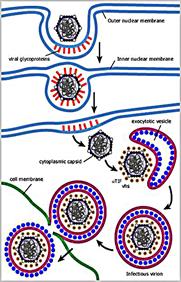
Проникнення вірусу в клітину відбувається услід за адсорбцією і включає "роздягання", тобто депротеїнезацію віріонів і зміну структури їх нуклеопротеїду. Проникнення віріона в клітину здійснюється у відповідність з тим же механізмом, що і проникнення в клітину позаклітинних поживних речовин, деяких гормонів, ростових факторів та ін., тобто шляхом рецепторного ендоцитоза. Механізм цього явища полягає в тому, що після прикріплення вірусів до клітинних рецепторів утворюються ендоцитарні вакуолі, які, зливаючись з іншими внутрішньоклітинними вакуолями, формують рецептосому - велику вакуоль з коротким періодом життя, що містить, пов'язану з рецепторами вірусну частку.
Рис. 20. Адсорбція віріонів вірусу вісповакцини на поверхні культури клітин нирки зеленої мавпи (х24000).
У рецептосомі відбувається взаємодія поверхневих білків віріонів з ліпідами стінки вакуолі, що призводить до злиття ліпопротеїнової оболонки (пеплоса) оболонкових вірусів з плазмолемою і виходу внутрішнього компонента вірусів в цитоплазму. Цей механізм проникнення в клітину є універсальним як для багатьох оболонкових, так і для без оболонкових вірусів, що належать до різних таксономічних груп.
А Б
Рис. 21. Послідовні етапи проникнення вірусу в клітини тварин і його "роздягання". А. Проникнення при частковому злитті оболонки вірусу і клітинної мембрани; Б. Проникнення вірусу з формуванням ендоцитозної вакуолі.
Взаємодія вірусної і клітинної мембран відбувається в більшості випадків при низьких (5,0 - 5,5) значеннях рН, що необхідно для придбання білками злиття необхідної конформації.
Проникнення вірусів може здійснюватися через злиття мембран вірусу і клітини. Злиття обумовлене наявністю відповідних вірусних глікопротеїнів, внаслідок чого внутрішні структури вірусів опиняються в цитоплазмі заражених клітин, а вірусні оболонки - на поверхні клітин.
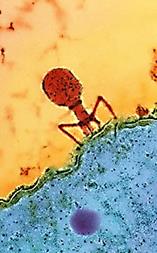
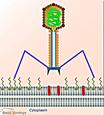
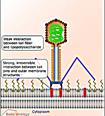
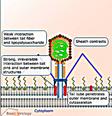
Інший шлях проникнення реалізується у випадку бактерій і вірусів бактерій - бактеріофагів. Цей шлях - проникнення в клітину тільки нуклеїнової кислоти, добре вивчений на моделях Т-парних (Т-2, Т-4 і так далі) фагів кишкової палички. Фагова ДНК потрапляє в клітину в результаті скорочення білкового чохла відростка вірусу (рис. 16), чому передує адсорбція віруса на клітинній стінці бактерії активація вірус-кодованого ферменту - лізоциму і часткове руйнування ним полісахаридного комплексу (рис. 22а, 22б).
1 2 3 4
Рис. 22а. Послідовні етапи інфікування бактеріальної клітини бактеріофагом групи Т-фагів кишкової палички
1. Адсорбція вірусу на клітинній поверхні;
2. Часткове руйнування полісахаридного комплексу клітинної стінки лізоцимом фага;
3. Введення білкової "голки" фага через мембрану клітини;
4. Інфікування клітини фаговою ДНК
Роздягання. Для придбання інфекційної активності вірусна частка повинна лишитися ліпопротеїдної оболонки і частково - білків капсиду. Роздягання вірусу - багатоетапний процес, причому у деяких вірусів останні етапи роздягання проходять в ядрі інфікованої клітини.

|
Рис. 22б. Проникнення ДНК фага Т2 в бактерію кишкової палички після адсорбції вірусу.
Пізні стадії. Інформація, записана у вірусному геномі, реалізується шляхом перемикання клітини на синтез вірусспецифічних молекул, що здійснюється двома способами:
1) синтезом коротких інформаційних РНК для індивідуальних білків (віруси герпесу, віспи, ВІЛ та ін.);
2) використанням вірусом власних геномів як інформаційних РНК з послідуючою трансляцією гігантських поліпептидів - попередників функціонально активних білків віруси поліомієлиту, гепатиту та ін.).
Перший спосіб характерний для вірусів ДНК-вмісних, а також для РНК- вмісних вірусів з негативним геномом і вірусів з фрагментованим геномом, у яких кожен фрагмент є окремим геном.
Другий спосіб - для РНК- вмісних вірусів з позитивним геномом.
У першому випадку інфекційний цикл в зараженій клітині починається з утворення іРНК, а в другому випадку - з синтезу білку. Поліпептид, що утворюється, - посередник розрізає клітинними і вірус-специфічними протеазами на функціонально активні. Треба вказати на те, що нарізання одного-двух білків-попередників або видалення невеликої ділянки поліпептиду відзначається при репродукції практично усіх вірусів і є необхідним для придбання ними інфекційної активності (цей процес називається протеолітичною активацією). У оболонкових РНК- вмісних вірусів нарізаються зазвичай поверхневі білки – глікопротеїди, що беруть участь в проникненні в клітину, тому протеолітична активація пов'язана із здатністю вірусу проникати в клітину.
Важливою особливістю розмноження вірусів є те, що компоненти вірусу синтезуються в клітині окремо і лише потім з'єднуються в зрілу вірусну частку (диз’юктивний спосіб розмноження). Синтез вірусних нуклеїнових кислот і вірусних білків може відбуватися не одночасно і в різних частинах клітини.
Передусім, після інфікування клітини вірусом настає різке пригнічення синтезу клітинних нуклеїнових кислот і білку. Подальші етапи пов'язані з процесами транскрипції і трансляції вірусної нуклеїнової кислоти.
Транскрипція.
Експресія вірусних геномів регулюється на рівні транскрипції, що призводить до утворення інформаційних РНК. Транскрипція вирусспецифічної інформації відбувається як за допомогою клітинних (більшість вірусів ДНК- вмісних), так і за допомогою вірусних ферментів-транскриптаз (віруси віспи, гепатиту В, грипу). Транскрипція РНК- вмісних вірусів здійснюється за участю транскриптаз, що кодуються вірусним геномом, які можуть бути як структурними, так і неструктурними білками. Наступна особливість полягає в тому, що синтетичний апарат еукаріотичних клітин не пристосований для трансляції поліцистронних іРНК.
Віруси обходять цю перешкоду або шляхом синтезу іРНК, що включає декілька генів і що кодує великий "поліпротеїн", який потім розрізає на індивідуальні білки, або шляхом розподілу генетичної інформації по фрагментах генома, кожен з яких є окремим геном і транскрибується в окрему іРНК.
У ДНК- вмісних вірусів іРНК синтезується в певній послідовності: спочатку транскрибуються надранні гени, потім ранні (ферменти, регулювальники - неструктурні білки), проміжні, і в останню чергу - пізні. Ферменти реплікації кодуються ранніми генами, структурні білки - пізніми.
У паповавірусів ранні і пізні гени транскрибуються в різних напрямах на різних нитках кільцевої дволанцюжкової ДНК. Багато генів під час транскрипції перекривають один одного. Деякі ділянки ДНК прочитуються в різних рамках зчитування, внаслідок чого можуть утворитися два різні білкові продукти. Ці особливості транскрипції сприяють значній економії генетичного матеріалу і збільшенню місткості генетичної інформації вірусу.
Для багатьох ДНК вірусів характерний синтез білков-трансактиваторів, що включають транскрипцію і стимулюють експресію генів. Характерною особливістю вірусних трансактиваторів є відсутність специфічності: вони здатні активувати як експресію генів інших вірусів, так і генів клітин еукаріот.
Одним із способів регуляції транскрипції є фосфорилювання і дефосфорилювання білків, що впливає на зв'язування білків з нуклеїновою кислотою.
Механізм реплікації забезпечує відтворення дочірніх вірусних геномів, які є точною копією материнських. Механізм реплікації у різних вірусів неоднаковий. У вірусів ДНК- вмісних в реплікації бере участь клітинна ДНК-полімераза.
Реплікація кільцевих двонитчастих ДНК починається з однієї точки і йде одночасно в обох напрямах з однаковою швидкістю. Як і у разі реплікації клітинної ДНК відбувається переривчастий синтез ініціаторів (приманок) - коротких ділянок РНК (РНК-праймерів) і коротких фрагментів ДНК (фрагменти Оказакі), які потім ковалентно зв'язуються в зростаючий ланцюжок. При утворенні лінійної дволанцюжкової ДНК (аденовіруси) вони синтезуються з обох кінців в напрямі від 5¢- до 3¢-кінця, використовуючи вірусспецифічну ДНК-полімеразу.
Реплікація вірусних РНК-геномів відбувається тільки за участю вірусспецифічних РНК-полімераз (тобто ферментів, синтез яких кодується вірусним геномом) і має потребу в утворенні антигенома - повнорозмірної дзеркальної копії комплементу вірусного генома:
РНК вірусна Þ РНК копія (антигеном) Þ РНК вірусна (дочірня)
¯ ¯
поліпротеїд РНК-поліимераза ¯
¯¯¯¯¯(процесинг) ®
Дата добавления: 2015-05-03; просмотров: 1760;