ОСОБЕННОСТИ ПЕРВОГО ЭТАПА ГБФ-ПУТИ
1. 1-Й ЭТАП ПОЛНОСТЬЮ ОБРАТИМ.
Из 10 стадий 1-го этапа 7 полностью обратимы. Для остальных 3-х этапов есть обходные пути.
Для гексокиназы (1-я стадия) обходной обратный путь катализируется глюкозо-6-фосфатазой.
Для фосфофруктокиназы (3-я стадия) обходной обратный путь катализируется фруктозодифосфатазой.
Третьим необратимым звеном 1-го этапа является 10-я стадия. В прямом направлении эта реакция катализируется пируваткиназой, а обходной обратный путь протекает через 2 стадии. В 1-й реакции СО2 присоединяется к ПВК с образованием щавелево-уксусной кислоты (ЩУК). Эта реакция катализируется пируваткарбоксилазой. Эта реакция синтеза требует затраты 1 АТФ. Кофермент пируваткарбоксилазы: биотин (vit H).
Вторая реакция: ЩУК поступает в цитоплазму (с участием челночных механизмов) и превращается в ФЕП. Эта реакция требует затраты ГТФ.
С учетом 7-й стадии, где происходит субстратное фосфорилирование, получается, что на пути от пирувата до глюкозы расходуется 3 АТФ, то есть 6 АТФ в расчете на каждую молекулу глюкозы, синтезированную из ПВК.
Если не считать НАДН2, который образуется на 6-й стадии, а при обратном пути расходуется, то распад глюкозы до ПВК требует расхода 6 АТФ. Это подтверждает, что на любой биосинтез определенного метаболита требуется энергии больше, чем образуется при распаде этого же метаболита.
ВТОРАЯ ОСОБЕННОСТЬ ПЕРВОГО ЭТАПА ГБФ-ПУТИ: НА 1-М ЭТАПЕ НЕТ ПОЛНОЙ ЗАВИСИМОСТИ ОТ КИСЛОРОДА.
Среди 10-ти стадий 1-го этапа только одна является окислительной.
Это 6-я стадия: ФГА ----> 1,3-бисфосфоглицериновая кислота. Сама эта реакция не требует кислорода. Обычно НАДН2 поступает в митохондрии и по дыхательной цепи отдает водород на кислород.
Транспорт водорода от цитоплазматического НАДН2 к митохондриальному НАДН2 обычно обеспечивается малат-аспартатным челночным механизмом, изображенном на рисунке.
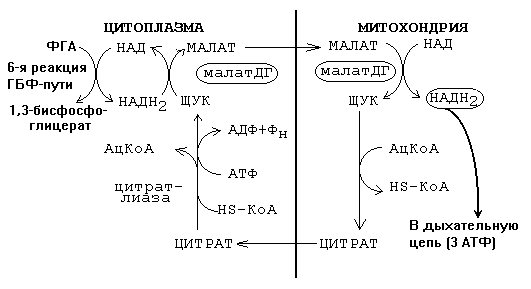
В этом случае весь 1-й этап работает как часть ГБФ-пути.
Дата добавления: 2015-02-28; просмотров: 925;