Регуляция генов у бактерий
Как и в предыдущих главах, начнем с простых биологических систем, то есть с бактерий, при изучении которых этот вопрос впервые был поставлен. Исследования велись преимущественно в 1950-х и 1960-х годах, в основном в парижском Институте Пастера; эксперименты проводили французские ученые Франсуа Жако, Жак Моно и некоторые американские специалисты, приехавшие в Париж для совместной работы. Первые опыты касались одной интересной особенности Е. coli, обитающей в кишечнике млекопитающих. Известно, что млекопитающие, особенно на первых порах жизни, потребляют много молока, а основной сахар молока — лактоза. Следовательно, в процессе долгой эволюции бактерии приспособились жить в среде с лактозой, и у бактерий Е. coli имеются ферменты для ее переработки. Но поскольку уровень лактозы в среде не всегда одинаков, то у бактерий должен существовать какой-то механизм для определения присутствия лактозы. Е. coli — хорошо адаптированный организм, и метаболизм лактозы в нем прекрасно отрегулирован. Лактоза представляет собой двойную молекулу сахара (дисахарид), состоящую из простых Сахаров — галактозы и глюкозы. На первой стадии метаболизма фермент в-галактозидаза расщепляет дисахарид на составные части, которые клетка может перерабатывать на последующих стадиях. Если выращивать Е. coli в среде без лактозы, то бактерии произведут небольшое количество в-галактозидазы. Если же добавить в культуру лактозу, то через 3—5 минут в клетках можно наблюдать существенные изменения: они начинают вырабатывать фермент в 1000 раз быстрее прежнего, и только один этот фермент может составить несколько процентов от общей массы бактерии. Стоит удалить лактозу (при помощи фильтров или центрифугирования), как тут же в течение нескольких минут производство ферментов снижается до первоначального уровня.
Для исследования этого механизма Моно с коллегами использовали ставший классическим метод мутационного анализа. Они отбирали мутантов, не способных перерабатывать лактозу, и обнаружили несколько мутантов с дефектной в-галактозидазой, которых назвали мутантами lacZ. Выяснилось, что эти мутанты производят нормальный фермент, но, тем не менее, не могут расти в среде с лактозой. У них оказался дефектный белок галактозидпер-меаза, который доставляет галактозу через клеточную мембрану внутрь клетки. Мутанты с дефектной галактозидпермеазой получили название lacY. Картирование показало, что гены lacZ и lacY, названные так по мутантам, располагаются рядом друг с другом.
У наиболее интересных мутантов наблюдался дефект в регуляторной системе, поэтому они не могли начинать или останавливать экспрессию генов lac. Мутанты, названные lacI, одновременно вырабатывали в-галактозидазу и пермеазу и не имели средств их контроля. Примечательно, что ген lacIрасположен рядом с генами Z и Y.
Последующие эксперименты позволили выяснить механизм контроля. Прежде всего, следует уяснить, что экспрессия гена подразумевает его транскрипцию — синтез матричной РНК.Вспомним, что транскрипцию осуществляет большой фермент РНК-полимераза, которая начинает транскрипцию с определенного места, примыкающего к кодирующему региону и называемого промотором. Для генов lacZ и lacY промотором служит небольшой участок между I и Z. Полимераза движется и производит РНК-транскрипт в определенном направлении: часто говорят, что чтение гена происходит сверху вниз. В таком случае промотор расположен выше гена lacZ. Ген lacI, который теперь называют геном-регулятором, кодирует белок lac-penpeccop. Это аллостеричес-кий белок (см. с. 281), который имеет два центра связывания и поэтому может присоединяться к двум разным молекулам. Один центр специфичен для небольшой последовательности ДНК, называемой оператором, которая располагается между промотором и геном lacZ. В отсутствие лактозы репрессор связывается с оператором, блокируя транскрипцию, поэтому гены Z и Yоказываются невыраженными.
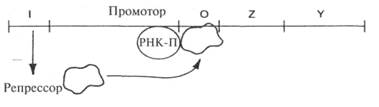
Кроме того, у белка-репрессора имеется центр связывания с лактозой, поэтому если в среде есть лактоза, она связывается с репрессором, вследствие чего репрессор слегка изменяет свою форму и уже не может связываться с оператором. Поэтому репрессор отсоединяется от оператора, позволяя осуществлять транскрипцию генов Z и Y.
Таким образом, гены Z и У выражаются совместно. Такие гены, контролируемые одним оператором, называются опероном.
Геном бактерий насчитывает многие виды оперонов, которые регулируются по-разному. Например, регуляция генов биосинтеза, кодирующих ферменты для производства таких клеточных компонентов, как аминокислоты, происходит по иной схеме, противоположной описанной. Предположим, что клетка находится в среде, богатой всеми необходимыми аминокислотами. Если регуляция генов клетки происходит правильно, то она должна прекратить тратить лишнюю энергию на производство избыточных материалов. Гены биосинтеза ферментов также образуют опероны, но их регулируют другие виды бел-ков-репрессоров, которые связываются с оператором (и тем самым блокируют транскрипцию генов) только при наличии избытка аминокислот. Например,
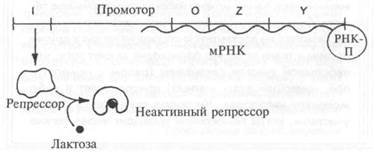
синтез гистидина кодируется большим блоком генов, который регулируется одним оператором и репрессором. Этот репрессор связывается с оператором и предотвращает транскрипцию генов только в том случае, когда в клетке имеется избыток гистидина. Если концентрация гистидина уменьшается, молекулы гистидина отсоединяются от молекул репрессора. Репрессор больше не может связываться с оператором, поэтому с оперона снова можно осуществлять транскрипцию.
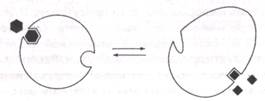
| Белки, которые связываются Мы много говорили о белках-ферментах, которые служат катализаторами и ускоряют химические реакции, но белки могут выполнять и другие функции в клетке. При изучении биологии важно уяснить, что многие белки временно или постоянно связываются с другими молекулами, называемыми лиганда-ми. Лигандом может быть другой белок, и тогда образуются комплексы белков, вроде тех, что поддерживают форму клетки. Наши красные кровяные клетки, например, имеют форму толстых вогнутых дисков, и эту их форму образуют несколько белков, постоянно связанных друг с другом и расположенных непосредственно под поверхностью клетки. Временное связывание можно наблюдать на примере гемоглобина — красного белка, наполняющего эти клетки. Гемоглобин доставляет кислород из легких в другие органы и ткани тела; это происходит за счет того, что небольшой участок связывания (рядом с гемогруп-пой, имеющей атом железа) присоединяет к себе молекулу кислорода. Кислород связывается с этим участком, когда гемоглобин проходит через легкие и отсоединяется в тех участках тканей, где мало кислорода. Одни из самых важных видов белка — белки-ре-цепторы. Их функция состоит в том, чтобы связываться со специфическими лигандами и, как правило, передавать информацию об их присутствии. У белка-рецептора имеется небольшой центр связывания, который по своей форме и химической структуре соответствует определенной молекуле. Рецепторы на языке и в полости носа захватывают молекулы из пищи или из воздуха, позволяя нам чувствовать вкус и запах. Нервная система состоит из многих нейронов с длинными отростками, которые соединяются между собой, как провода в телефонной системе. Каждая клетка посылает сигнал соседней, выделяя сигнальные молекулы, которые связываются с рецепторами соседних нейронов и стимулируют их пересылать сигнал дальше. Точно так же и бактерии имеют рецепторы, способные распознавать те или иные аминокислоты. Когда рецепторы бактерий обнаруживают сахара или аминокислоты, они посылают сигналы клетке, и та передвигается поближе к источнику питательных веществ. Все эти белки выполняют функции благодаря своей подвижности и гибкости. Связываясь с лигандами, они слегка меняют свою форму, что позволяет им выполнять следующую функцию — стимулировать аппарат передвижения бактерии или генерировать электрический сигнал в нейроне. Когда молекула гемоглобина соединена с молекулой кислорода, ее форма слегка отличается от молекулы гемоглобина без кислорода. Способность менять форму имеет особое значение для аллостерических белков; у таких белков имеется два центра связывания, соответствующих двум разным лигандам, и форма белка зави Продолжение на след. странице |
| сит от того, какой именно лиганд связан с ним в данный момент. В классическом случае аллостеричес-кий белок имеет активный центр для своего субстрата и вторичный центр для регуляторного лиганда: |
|
| Такого рода белки часто стоят первыми в метаболической цепи и регулируют метаболический путь посредством обратной связи. Регуляторный лиганд в данном случае является конечным продуктом метаболического пути, и при своем избытке в клетке он связывается с молекулами-ферментами, слегка изменяя их и предотвращая их активность. Затем, когда уровень концентрации продукта падает, его молекулы отсоединяются от ферментов, и ферменты возобновляют свою активность. Благодаря такому механизму клетка получает достаточное количество необходимых ей компонентов, но не тратит энергию на производство лишних веществ. |
Дата добавления: 2015-02-28; просмотров: 909;