Окисление органических веществ в клетках, сопровождающееся потреблением кислорода и синтезом воды, называется тканевым дыханием.
Цепь переноса электронов от окисляемых органических субстратов к кислороду, называется дыхательной цепью (рис. 19).
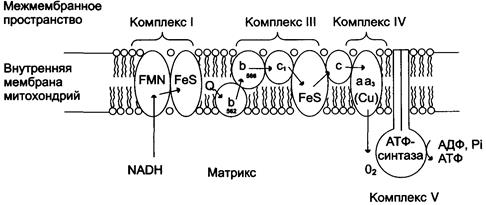
Рис. 19.Митохондриальная цепь переноса электронов.Комплекс I содержит FMN и не менее пяти железо-серных белков(FeS). Комплекс III включает две разные формы цитохрома b (с максимумами поглощения 562 и 566, один FeS-белок и цитохром с. Комплекс IV содержит цитохромы а1 и а3 и два иона меди. Комплекс II (сукцинатдегидрогеназа) на рисунке не показан. Комплекс V - АТФ-синтаза (Авдеева Л.В., Павлова Н.А., Рубцова Г.В., 2005)
Перенос электронов от окисляемых субстратов к кислороду происходит в несколько этапов. В нем участвует большое количество промежуточных переносчиков, каждый из которых способен присоединять электроны от предыдущего компонента и передавать следующему. За исключением убихинона (кофермент Q) и цитохрома С промежуточные переносчики кислорода – сложные белковые комплексы. Основные переносчики электронов встроены во внутреннюю мембрану митохондрий и организованы в 4 комплекса, расположенных в определенной последовательности (векторность). Среди них: НАДH-дегидрогеназа (НАДH-Q-редуктаза) – комплекс I; QH2-дегидрогеназа (коэнзим Q-цитохром с-редуктаза) – комплекс III; цитохромоксидаза – комплекс IV; АТФ-синтаза – комплекс V. Каждое из этих звеньев дыхательной цепи специфично в отношении донора и акцептора электронов. В этой последовательности окислительно-восстановительные потенциалы по мере приближения к кислороду становятся все более положительными.
Таким образом, электроны, поступающие в дыхательную цепь, по мере передвижения от одного переносчика к другому по возрастающему редокс-потенциалу, теряют (высвобождают) свободную энергию. Часть этой энергии рассеивается в виде тепла, а часть (40-45%) аккумулируется в молекулах АТФ.
Снижение свободной энергии происходит на каждом этапе цепи переноса электронов, т.е. энергия высвобождается порциями. Принципиальное положение заключается в том, что процесс переноса электронов в дыхательной цепи и синтез АТФ энергетически сопряжены. При этом перенос электронов на кислород сопровождается высвобождением свободной энергии (экзергонический процесс), а фосфорилирование АДФ (синтез АТФ) – запасанием (притоком) свободной энергии (эндергонический процесс). Синтез АТФ из АДФ и Н3РО4, осуществляемый за счет энергии переноса электронов, называется окислительным фосфорилированием. Этот процесс протекает во внутренней мембране митохондрий.
Механизм сопряжения транспорта электронов в дыхательной цепи и окислительного фосфорилирования сегодня обосновывается с позиций хемиосмотической гипотезы (Митчелл, 1961). В соответствии с этой гипотезой часть энергии, высвобождаемой при транспорте электронов, расходуется на выкачивание протонов из матрикса митохондрий в межмембранное пространство. В результате между двумя сторонами внутренней митохондриальной мембраны (непроницаемой для протонов) создается концентрационный и электрический градиенты, обусловленные их скоплением на внешней поверхности внутренней мембраны. В совокупности эти градиенты формируют электрохимический потенциал ΔµН+.
Наиболее активный транспорт протонов в межмембранное пространство, необходимый для формирования ΔµН+ происходит на участках дыхательной цепи, соответствующих расположению комплексов I, III и IV. Эти участки называют пунктами сопряжения дыхания и фосфорилирования. Механизм транспорта протонов в пунктах сопряжения через митохондриальную мембрану недостаточно ясен, однако важная роль в этом процессе отводится коэнзиму Q. Предполагается, что когда ионы Н+ возвращаются из цитозоля межмембранного простанства в матрикс, проходя через ионные каналы АТФ-синтазы, именно этот градиент служит движущей силой для синтеза АТФ.
Эффективность синтеза молекул АТФ оценивается коэффициентом окислительного фосфорилирования, представляющего собой отношение количества фосфорной кислоты, использованной на фосфорилирование АДФ, к атому кислорода, поглощенного в процессе дыхания (Р/О).Для НАДН теоретический минимум Р/О равен 3; для сукцината – 2. Фактически эти величины ниже.
Окисление субстратов и фосфорилирование АДФ в митохондриях прочно сопряжены. Скорость использования АТФ регулирует скорость потока электронов в дыхательной цепи. Если АТФ не используется и его концентрация в клетке возрастает, поток электронов к кислороду прекращается. Рост АДФ, наоборот, увеличивает окисление субстратов и поглощение кислорода. Зависимость интенсивности дыхания митохондрий от концентрации АДФ называется дыхательным контролем. Этот контроль обеспечивает соответствие скорости синтеза АТФ к потребности клетки в энергии. Благодаря такому контролю относительная концентрация АТФ/АДФ в клетке изменяется в узких пределах, тогда как потребление клеткой энергии, т.е. скорость оборота АТФ/АДФ может меняться в десятки раз. За сутки у человека синтезируется 40-60 кг АТФ и столько же распадается. Продолжительность жизни каждой молекулы АТФ составляет около 1 минуты. Общее содержание АТФ в организме – 40-50 г.
Дата добавления: 2015-02-05; просмотров: 3266;